摘 要:以伊犁河谷的大麻(Cannabissativa)为研究对象,分析林地、玉米地、麦地、路边等4种样区大麻根、茎、叶生态化学计量特征,了解植物生长机制,研究生境和器官对大麻生态化学计量特征的影响,阐明大麻生长过程中的限制性元素。研究结果表明,在林地、路边大麻各器官的碳含量以及林地大麻C∶P表现为茎>根>叶,玉米地中大麻碳含量为茎>叶>根,在麦地为根>叶>茎,各样区大麻氮含量和磷含量及林地、麦地大麻N∶P表现为叶>茎>根,各样区大麻C∶N及玉米地、麦地、路边大麻C∶P表现为根>茎>叶,在玉米地、路边的大麻N:P为叶>根>茎。不同生境大麻不同器官生态化学计量特征存在一定差异性,路边大麻叶、根的碳含量极显著高于其他样区(P<0.01),茎的碳含量显著高于麦地(P<0.05),且高于其他样区,但不显著(P>0.05);玉米地大麻叶、茎的磷含量和C:N显著高于其他样区(P<0.05),根的磷含量高于其他样区,但林地与玉米地不显著(P>0.05),玉米各器官氮含量、C∶P和N∶P极显著低于其他样区(P<0.01),其中玉米地与麦地大麻茎的氮含量差异性不显著(P>0.05)。各生境大麻叶的N∶P都小于14,说明大麻生长受氮素限制。C∶N、C∶P都与相应的氮含量、磷含量表现为极显著(P<0.01)的负相关关系,氮含量与磷含量表现为极显著(P<0.01)的正相关关系,表明大麻氮、磷元素需求变化一致。通过GLM(GeneralLinearModel)分析,样区对碳含量的影响较大,器官对氮含量和磷含量及C∶N、C∶P、N∶P的影响较大。
关键词:大麻,生境,器官,生态化学计量特征,伊犁河谷
生态化学计量学这一概念首先是由Elser等明确提出[1−2]。现如今,生态化学计量学已经是生态学研究的热点问题,其理论最早应用于水生生态系统的研究中,但在近20年以来,生态化学计量学的研究领域已逐渐扩展至陆地生态系统中,尤其在陆生植物的氮、磷化学计量学方面得到逐步的发展,陆地植物生态化学计量特征主要是碳、氮、磷的计量关系[3]。通过对植物生态化学计量特征的研究,可了解植物生长过程中养分利用情况。在曾冬萍等的研究中,个体水平的生态化学计量特征表明植物叶、茎、根存在关联性[4]。王振南等对植物生态化学计量与非生物因子响应的研究表明,植物碳、氮、磷含量及其生态化学计量特征体现了环境因子与植物生态化学计量特征的耦合关系,反映了植物对环境变化的响应和适应[5]。因此,通过对植物碳、氮、磷生态化学计量学特征的研究,有助于理解植物的生长机制,探讨植物器官与生境对植物的影响。
大麻(Cannabissativa),桑科大麻属植物,中国各地有栽培或野生,新疆常见野生,本次研究对象为伊犁河谷野生大麻。大麻为一年生或多年生直立草本植物或亚灌木,雌雄异株,含同株,大麻可广泛分布,适应性强,野生大麻可易见于抛荒地、沙滩、林缘、沟边及草甸处[6]。野生大麻繁殖力强,在适宜的环境条件下会排斥其他植物并扩大自身生长区域,在棕黑土或黄壤土大量分布,与栽培种类类似但茎较细长、分支少、籽粒小[7]。大麻有一定药用价值,也可用来织麻布或纺线,制绳索、造纸等获得经济效益。现如今对大麻的研究主要为大麻的选育栽培及其药理作用研究,野生大麻多为研究其遗传多样性,为大麻栽培杂交以及工业大麻品种改良提供参考依据[6,8],对大麻的生态化学计量特征研究鲜见报道。
为进一步解析大麻器官中碳、氮、磷含量及其化学计量特征以及不同生境下大麻的生态化学计量特征,阐明大麻的生长机制,通过对伊犁河谷中林地、玉米地、麦地、路边等4个样区内的大麻进行采样,测量大麻叶、根、茎不同器官中的碳、氮、磷含量,分析器官与生境对大麻整体生态化学计量特征的影响,为大麻的生长及其及管控提供一定的理论依据。
1材料与方法
1.1研究区概况
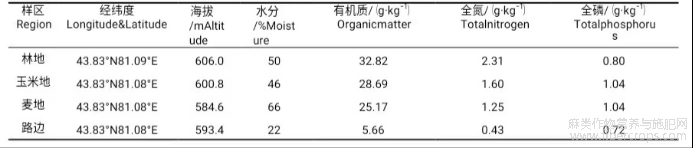
研究区雀儿盘村位于伊犁河谷察布查尔锡伯自治县。察布察尔锡伯自治县的地理位置位于北纬43°17′—43°57′,东经80°31′—81°43′,在新疆原伊犁地区板块的西部。察布查尔县属于大陆性北温带温和干旱气候,全年有效光照时数达2846h,无霜期177d,积温3800℃,年均气温7.9℃,年平均降水量222mm[9−10]。本文选取了林地、玉米地、麦地、路边等4个生境(表1)。
表1 不同样区土壤理化因子特征
1.2样品采集
将样区设置在伊犁河谷察布查尔县雀儿盘村,采样时间为2019年7月,依次将4个生境(林地、玉米地、麦地、路边)布设为样区Ⅰ、样区Ⅱ、样区Ⅲ、样区Ⅳ。4个样区的土壤理化性质不同,水分含量样区Ⅲ>样区Ⅰ>样区Ⅱ>样区Ⅳ;有机质含量、全氮含量以及全磷含量皆为样区Ⅰ>样区Ⅱ>样区Ⅲ>样区Ⅳ。每个样区随机设置3个1m×1m的样方,样方间距为10m。在每个样方内采集大麻地上部分,然后将地面枯落物清理干净,每隔10cm采集1个植物根系样品(包括根茎和须根),采集深度20cm,采集表面根茎样品,共采集24份植物根系样本以及12个植物地上部分样品[11−13]。
1.3实验方法
土壤样品采集后进行风干,剔除样品中的植物部分和石块,研磨过100目筛备用。其中土壤的理化性质选取土壤水分、有机质、全氮、全磷。植物样品主要为野外采集的大麻样品带回实验室后洗净去除杂质,烘干后粉碎过筛,测定植物样品的有机质、全氮、全磷。
土壤水分采用烘干法测定;土壤以及植物各器官有机碳采用重铬酸钾容量-外加热法测定;全氮采用高氯酸-硫酸消化法测定;全磷采用酸溶-钼锑抗比色法测定,以上指标均采用鲍士旦的土壤农化分析方法[14]。C、N、P含量测定结果以单位质量的养分含量表示(g·kg−1),C∶N、C∶P、N∶P为质量比。
1.4数据处理
使用软件Microsoft Excel 2018对大麻各样区内不同器官的C、N、P测定数值进行统计并作图。用SPSS 20.0软件对以上数据进行分析:采用单因素方差分析检验4种环境下大麻根茎叶C、N、P的化学计量特征和土壤理化因子的差异,并采用Duncan法进行多重比较,生境和器官对大麻C、N、P含量及其化学计量比的影响采用GLM(General Linear Model)模型分析主因子效应和交互作用,分析每个因子的影响。
2结果与讨论
2.1不同样区大麻根茎叶碳、氮、磷含量
不同样区大麻根茎叶的碳、氮、磷含量存在差异(图1)。除了麦地中大麻的各器官有机碳含量差异性不显著,其余样区大麻各器官有机碳含量差异性极显著,麦地中茎的碳含量平均值为370.23g·kg−1,低于其他器官中碳含量,而其余样区中茎的碳含量都显著高于其他器官。林地与玉米地叶的碳含量差异性不显著,其余样区叶的碳含量差异性极显著;林地与麦地茎的碳含量差异显著,麦地与路边茎的碳含量差异显著;不同样区根的碳含量差异性皆极显著。
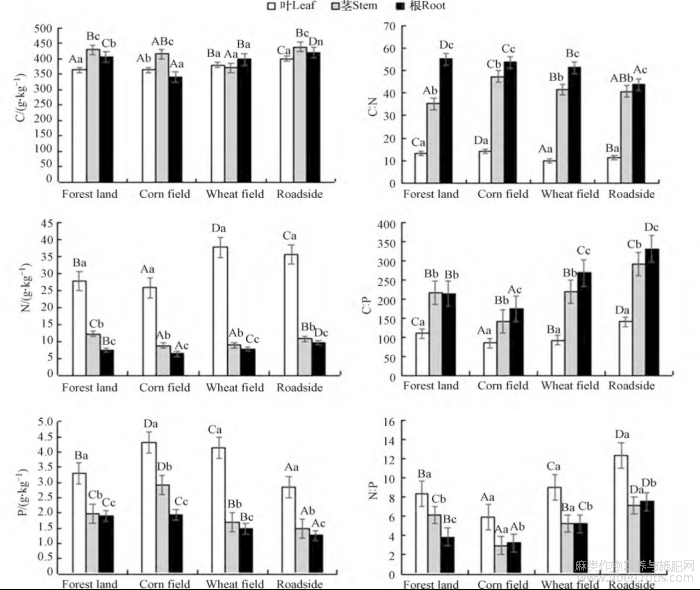
图1 不同样区大麻各器官C、N、P元素含量及其化学计量比
大写字母代表各器官不同样区的显著差异(P<0.05);小写字母代表各样区不同器官的显著差异(P<0.05);
各样区大麻氮含量为叶>茎>根,各器官氮含量的差异性极显著。不同样区中叶的氮含量差异性极显著,呈现为麦地>路边>林地>玉米地;玉米地与麦地中大麻茎的氮含量差异不显著,其余样区大麻茎的氮含量差异极显著;不同样区中根的氮含量差异性极显著,呈现为路边>麦地>林地>玉米地。
各样区大麻磷含量为叶>茎>根,林地中叶的磷含量与根茎的磷含量差异性极显著,其余样区各器官磷含量的差异性极显著。大麻叶的磷含量在玉米地与麦地差异性显著,其余样区差异性极显著;不同样区大麻茎的磷含量差异性极显著;林地与玉米地中根的磷含量差异性不显著,其余差异性极显著,大麻的叶、茎、根的磷含量皆是玉米地高于其他样区,玉米地大麻叶、茎、根磷含量均值分别为4.32、2.93、1.95g·kg−1。
2.2不同样区大麻根茎叶碳、氮、磷化学计量比特征
不同样区大麻根茎叶的C∶N、C∶P以及N∶P的化学计量比存在一定的变化规律(图1)。各样区大麻的C∶N为根>茎>叶,其中林地、玉米地、麦地大麻根的碳氮比均值为53.365,路边较小为43.707,麦地中大麻的茎和根C∶N差异性显著,其余器官的C∶N差异性极显著,林地、玉米地、路边中大麻各器官的C∶N差异性极显著。不同样区大麻叶的C∶N差异性极显著,呈现为玉米地>林地>路边>麦地;林地与玉米地茎的C∶N差异性极显著,林地、麦地与路边茎的C∶N差异性不显著,玉米地茎的C∶N高于其余样区,平均值为47.35;不同样区大麻根的C∶N差异性极显著,呈现林地>玉米地>麦地>路边。
林地大麻的C∶P为茎>根>叶,茎与根的C∶P差异性不显著,叶与茎和根的C∶P差异性极显著;其余样区的C∶P呈现为根>茎>叶,其中路边大麻根的碳磷比最高为331.067,玉米地、路边中各器官C∶P差异性极显著,麦地中茎与根的C∶P差异性显著,叶与茎和根的C∶P差异性极显著。大麻叶和根的C∶P在不同样区间差异性极显著,大麻茎的C∶P在林地与麦地之间差异性不显著,叶的C∶P为路边>林地>麦地>玉米地,根和茎的C∶P为路边>麦地>林地>玉米地。
林地、麦地大麻的N∶P为叶>茎>根,其中麦地大麻叶的氮磷比最大为9.066,林地中各器官的N∶P差异性极显著,麦地中根和茎的N∶P差异性不显著;玉米地、路边中大麻的N∶P为叶>根>茎,两个样区皆为根和茎的N∶P差异性不显著。大麻根和叶的N∶P为路边>麦地>林地>玉米地,茎的N∶P呈现为路边>林地>麦地>玉米地,各器官不同样区间的N∶P差异性极显著。路边大麻叶的氮磷比最高,为12.410。
2.3大麻元素含量与化学计量比的相关关系
相关分析表现(图2),不同样区大麻的C∶N、C∶P都与相应的氮含量、磷含量表现为极显著的负相关关系,以对数方程呈现。各样区大麻氮含量与磷含量表现为极显著的正相关关系,图2中线性回归方程能更具体反映其趋势。
2.4器官和样区对碳、氮、磷含量及化学计量比的影响
通过SPSS中的GLM分析,量化样区和器官对大麻整体碳、氮、磷含量及其化学计量比的影响。大麻的碳、氮、磷含量及其化学计量比受器官和样区各主效应以及两因素交互效应的影响程度不同(表2)。样区和器官对大麻碳含量的影响均为极显著水平,样区对碳含量的影响更大。样区和器官对大麻氮含量的影响均为极显著水平,其中器官对氮含量的影响较大。样区和器官对大麻磷含量的影响皆为极显著水平,其中器官对磷含量的影响更大。样区和器官对大麻的C∶N、C∶P、N∶P的影响皆达极显著水平,皆为器官影响较大。样区和器官的交互作用对碳、氮、磷含量及其比值的影响均为极显著性差异。
2.5大麻根茎叶碳、氮、磷含量及化学计量比的变化
植物器官中碳、氮、磷含量及其化学计量比会受器官组织和功能的影响而变化显著[15]。碳元素是大气是主要组成部分,也是植物干物质的主要元素。叶是植物光合作用的重要器官,是合成有机物的主要场所,是植物吸收、储存营养的主要地上器官,也是对环境变化反应最为敏感的器官[5,11,16−27]。叶通过光合作用,糖类物质在叶片中产生并积累,使得碳含量得到增长[11]。刘青海[16]的研究发现,大麻在生长过程中,叶片中的有机物占全株的比例是随着时间变化而呈现逐步下降的趋势。本研究中大麻采样时间已达到大麻生长发育期间的工艺成熟期,此时叶片中的有机碳含量占比已下降,因此不同生境下测出的大麻叶片中所含有的碳含量都会低于茎、根。茎是支持叶、果实等地上构件并连接根等地下构件的器官,其作用在于将水分、有机物、无机物等营养运输到植物的各个部分[16−27]。大麻属于草本植物,茎也可进行光合作用,同时叶片中通过光合作用产生的有机物通过茎运输到根部,在这个过程中茎的碳含量也得到了积累而增加。茎的碳含量占比是随着麻龄的增长而逐渐上升,到成熟期能增长到占全株的一半以上[16],故在本次研究中大麻的碳含量是茎中的含量大于叶和根。根是吸收、转运土壤中营养物质的主要器官,其主要功能是运输水分和养分[16−27],且根不能发生光合作用,因此根对碳含量的积累较少。大麻根部的碳含量占比相对于其他器官来说是最小的,其有机物的积累比较平稳,随着大麻的生长,所占比例会逐步减少[16],但本次研究中大麻根的碳含量在不同生境下并不是所有器官中最小的。
氮和磷是植物生长发育的重要物质,也是影响植物生长、分布的限制生长因子[4,17]。氮元素是构成蛋白质的主要成分,对植物的生长发育起着重要作用,影响着植物的产量。本次研究中,在不同生境下大麻氮含量在器官内变化趋势为:叶>茎>根,与游冲、朱秋丽、李从娟等[18−19,15]研究结果相同。叶片和根对氮的吸收是随着麻龄增长而增加,到成熟期氮的积累量达到最大;茎的氮元素随着麻龄的增长而增长直至到稳长期达到最大,到成熟期是时减少,成熟期对氮的吸收较少,氮的积累主要在叶和根,而茎部会出现氮元素倒流现象[16]。磷元素是核酸的重要组成元素,对植物的生长代谢以及对外界的适应能力都有着较大影响,植物体主要是通过土壤中磷来获取自身所需的磷元素。大麻对磷的吸收较少,呈现为前期高,后期低,在成熟期时吸收强度会稍有增强,叶和茎对磷的吸收积累会符合植株整体对磷的吸收趋势,而根部随磷的吸收会在后期逐渐减少[16],本文中磷含量的变化趋势是叶>茎>根,与上面所述一致。
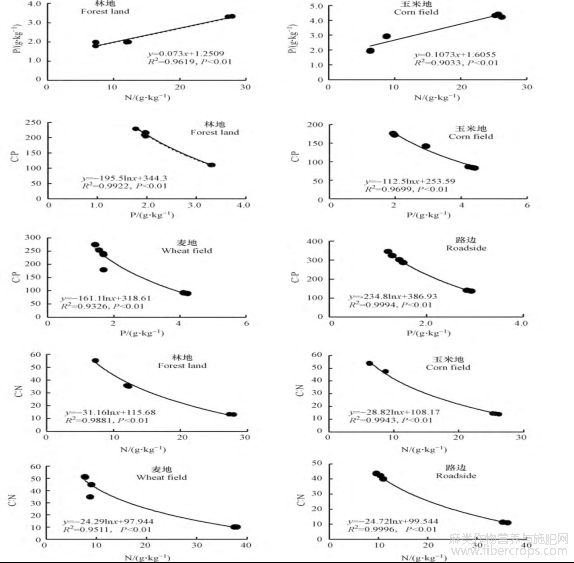
图2 不同样区大麻C∶N与N、C∶P与P及N与P之间的相关关系
植物在生长发育的过程中会通过调整自身以及各器官的营养比例来适应环境,C∶N、C∶P和N∶P这些化学计量比的变化则可体现这样的比例变化[28]。C∶N、C∶P在一定程度上可以反映植物吸收和同化碳元素的能力,表明了植物对营养元素的利用效率[19−20]。Sterner和Elser认为,生物的碳N∶P值与生长速率有很强的关系,在此基础上产生了“生长率假说”,即生物体的快速生长需要大量核糖体合成蛋白质,而核糖体中含有大量的磷元素,故生长速率较快的有机体通常有较低的C∶N和C∶P[1,4,29]。在本次研究中,不同生境下的C∶N、C∶P大体变化趋势皆为根>茎>叶,与罗艳等[11−12]研究结果相同,说明叶的生长速率相较于茎和根较快,故叶片的C∶N、C∶P最小,根最大。在孙连伟等[4]
研究说明,陆地植物体碳元素的含量通常维持在50%左右,因此陆地植物的生态化学计量学研究更偏向于氮和磷的化学计量特征。本次研究中叶的N∶P范围在5.959—12.410之间,根据Koerselman的研究表明,当植物叶片的N∶P小于14时表示植物生长很大程度受氮元素的限制[3,30],故本次研究中氮元素为主要限制生长因子。在本次研究中,N∶P的变化趋势呈现明显的叶显著高于根和茎,这与吴统贵[21]的研究中草本植物叶中N∶P高于根的发现一致。
表2 器官与样区对大麻C、N、P元素含量及其化学计量特征影响的GLM分析
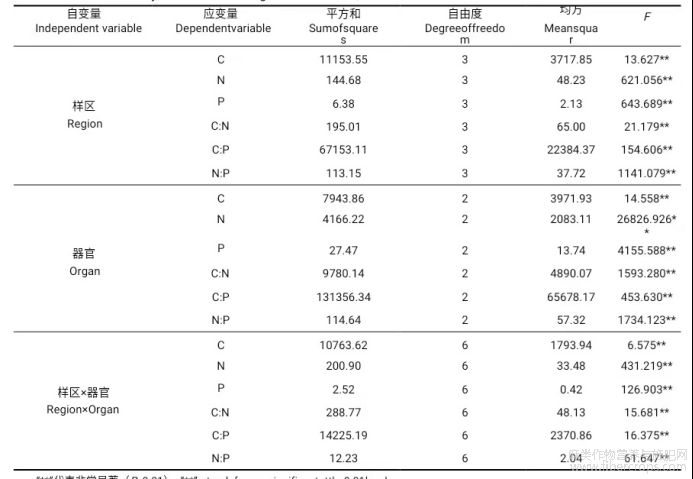
“**”代表非常显著(P<0.01).
2.6不同生境大麻碳、氮、磷含量及其化学计量比的变化
生长环境是影响植物生长的主要因素之一,植物的分布与生长状况受环境影响存在显著差异,植物体内不同器官的碳、氮、磷含量及其化学计量特征体现了植物适应环境变化的本质[28]。根据国家第二次土壤普查制定的土壤肥力分级标准,大麻的4个生境速效磷>40mg·g−1,为1级;林地的有机质在30—40mg·g−1,为2级,全氮>2mg·g−1,为1级;玉米地的有机质在20—30mg·g−1,为3级,全氮在1.5—2mg·g−1,为2级;麦地的有机质在20—30mg·g−1,全氮在1—1.5mg·g−1,为3级;路边的有机质<6mg·g−1,全氮<0.5mg·g−1,为6级。这说明大麻的4个生境磷含量比较充裕,其余营养元素含量差异较大。不同生境中大麻各器官的磷含量变化较为符合土壤全磷的变化,土壤氮含量的变化与大麻各器官中氮的变化不相一致。这与姜沛沛等[22]的研究结果较为符合,植物的氮、磷含量与土壤的氮、磷含量没有显著相关性。土壤水分对植物的影响会使得植物碳、氮、磷含量及其化学计量比产生差异[23]。在本次研究中,各器官碳含量最大的大麻在路边中,而路边中的水分是4个生境中最小,这与一些研究结论相同,有研究证实,植物叶片碳含量的变化会受降水量或人工灌溉量影响,表现为水分增加碳含量降低,而氮、磷含量会随着降水量或人工灌溉的增加而增加或变化不明显[5]。但也考虑由于是路边,相较于其他生境中的大麻,无其他植物遮挡,所获得的光照较强,使得植物的光合速率增大,有机碳的含量增加。王振南等[5]研究表明,不同的光照条件会影响植物光合作用和固碳、固氮相关酶的活性,从而影响植物碳、氮含量。
植物的生态化学计量特征受土壤中氮、磷的含量影响并随之变化[5]。在本次研究中生长在低营养环境路边中的大麻,C∶P、N∶P反而会比生长在其他高营养环境生境中的大麻高,这与罗艳等[11]研究结果相似,C∶P、N∶P可反应植物生长速率,高的C∶P和N∶P表征植物生长的速率较缓,在低营养环境下,植物生长将会减缓,植物对营养利用的效率将提高,C∶P将会增加。玉米地中的大麻C∶P最小,说明相较于林地、麦地和路边,玉米地中的大麻生长较快,扩展和影响农作物的速率较快,应该重点防治。植物叶片N∶P可作为一种指标来判断环境对植物生长养分供应,N∶P值越大说明氮充裕而磷相对不足,本次研究中氮元素是主要限制因子,所以各生境的大麻生长均受氮素限制。在高氮低磷的生境中,大麻将更多的磷元素分配到根部,根的氮磷值小,说明在高氮低磷的环境下大麻的根系生长速度快,增加了大麻地下竞争力;在高氮高磷和低氮低磷的生境下,大麻将磷元素更多地分配到了茎,茎的氮磷值小,说明高氮高磷和低氮低磷的环境下,大麻的地上竞争力较大。说明大麻在不同资源环境下对氮元素和磷元素吸收、利用以及分配不同,使得植物地上、地下的竞争优势不同,这与鬼针草的研究结果相似[24]。
2.7器官与生境对大麻碳、氮、磷元素含量及化学计量比的影响
大麻的生长发育,根据自身器官结构功能和生长环境的不同,碳、氮、磷含量及其化学计量比的分布存在差异。大麻通过叶、茎进行光合作用获取碳元素,各生境环境不同,光照条件不同,光合作用受光照条件的影响,故样区对大麻碳含量的影响较大。氮元素和磷元素对植物生长发育有着重要影响,主要是通过植物的根从土壤中获取,是蛋白质、核酸的主要元素。本次研究中各生境的大麻的生长都是受到氮素的限制,在这个生长条件下,大麻根据自身生长,调节器官内氮、磷的含量及其化学计量比,以适应不同生存条件。对鬼针草、黄顶菊的研究中也表明植物会通过调节自生养分含量及其计量比的变化以适应环境,增强自身的竞争优势[20,24]。在李从娟等[15]研究中,植物自身的遗传特性是决定植物叶片氮、磷含量的主要因素。有研究表明,植物的氮、磷含量与土壤的氮、磷含量没有显著相关性[22]。所以器官对氮、磷含量、C∶N、N∶P、C∶N的影响较大。
3结论
在林地、路边大麻各器官的碳含量以及林地大麻C∶P表现为茎>根>叶,玉米地中大麻碳含量为茎>叶>根,在麦地为根>叶>茎,各样区大麻氮含量和磷含量及林地、麦地大麻N∶P表现为叶>茎>根,各样区大麻C∶N及玉米地、麦地、路边大麻C∶P表现为根>茎>叶,在玉米地、路边的大麻N∶P为叶>根>茎。
不同生境大麻不同器官生态化学计量特征存在一定差异性,路边大麻叶、根的碳含量极显著高于其他样区,茎的碳含量显著高于麦地,且高于其他样区,但不显著;玉米地大麻叶、茎的磷含量和C∶N显著高于其他样区,根的磷含量高于其他样区,但林地与玉米地不显著,各器官氮含量、C∶P和N∶P极显著低于其他样区,其中玉米地与麦地大麻茎的氮含量差异性不显著。
各生境大麻叶的N∶P都小于14,说明大麻生长受氮素限制。对于玉米地中的野生大麻,由于其生长速率较快,应当注意防治,以减少大麻对农作物的影响。
不同样区大麻的C∶N、C∶P都与相应的氮含量、磷含量表现为极显著的负相关关系;不同样区大麻氮含量与磷含量表现为极显著的正相关关系。
样区和器官对大麻碳、氮、磷含量及其化学计量比值都有影响,样区对碳含量的影响较大,器官对氮含量和磷含量及C∶N、C∶P、N∶P的影响较大。
参考文献
[1]ELSERJJ,STERNERRW,GOROKHOVAE,etal.Biologicalstoichiometryfromgenestoecosystems[J].EcologyLetters,2000,3(6):540-550.
[2 ]ELSERJJ.Ecologicalstoichiometry:Fromseatolaketoland[J]. TrendsinEcology&Evolution,2000, 15(10):393-394.
[3]孙连伟,陈静文,邓琦.全球变化背景下陆地植物N/P生态化学计量学研究进展[J].热带亚热带植物学报,2019,27(5):534-540.
[4]曾冬萍,蒋利玲,曾从盛,等.生态化学计量学特征及其应用研究进展[J].生态学报,2013,33(18):5484-5492.
[5]王振南,杨惠敏.植物碳氮磷生态化学计量对非生物因子的响应[J].草业科学,2013,30(6):927-934.
[6]杨柳秀,李超然,高雯.大麻化学成分及其种属差异研究进展[J].中国中药杂志,2020,45(15):3556-3564.
[7]汤志成.野生大麻种质资源遗传多样性分析[D].昆明:云南农业大学,2013:1-47.
[8]汤志成,陈璇,张庆滢,等.野生大麻种质资源表型及其RAPD遗传多样性分析[J].西部林业科学,2013,42(3):61-66.
[9]赛牙热木·哈力甫,宋瑞清,艾克拜尔·伊拉洪,等.察布查尔县土壤碳氮磷钾垂直分布规律研究[J].干旱区地理,2018,41(3):582-591.
[10]徐莉,闫俊杰,陈晨,等.新疆察布查尔县农耕层土壤养分空间特征及其与地形因子的关系[J].水土保持通报,2018,38(4):38-45.
[11]罗艳,贡璐.塔里木盆地南缘不同生境下芦苇生态化学计量特征[J].生态学杂志,2016,35(3):684-691.
[12]罗艳,贡璐,李杨梅.塔里木河上游绿洲农田不同生育期玉米根茎叶生态化学计量特征[J].水土保持研究,2018,25(2):112-119.
[13]崔东,陈亚宁,李卫红,等.伊犁河谷苦豆子C、N、P含量变化及化学计量特征[J].生态学报,2018,38(17):6163-6170.
[14]鲍士旦.土壤农化分析(第三版)[M].北京:中国农业出版社,2000:1-495.
[15]李从娟,徐新文,孙永强,等.不同生境下三种荒漠植物叶片及土壤C、N、P的化学计量特征[J].干旱区地理,2014,37(5):996-1004.
[16]刘青海.大麻对氮、磷、钾营养元素吸收特性的研究[J].中国麻作,1983(4):36-41.
[17]万芳,蒙仲举,党晓宏.荒漠草原建群种及其枯落物的C、N、P生态化学计量特征[J].东北林业大学学报,2020(2):29-33.W
[18]游冲.滨海不同生境盐地碱蓬的化学计量关系与代谢调节策略[D].天津:天津理工大学,2019:1-62.
[19]朱秋丽.闽浙沪滨海湿地草本植物碳氮磷生态化学计量特征[D].福建:福建师范大学,2017:1-82.
[20]屠臣阳,皇甫超河,姜娜,等.不同生境黄顶菊碳氮磷化学计量特征[J].中国农学通报,2013,29(17):171-176.
[21]吴统贵,吴明,刘丽,等.杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化[J].植物生态学报,2010,34(1):23-28.
[22]姜沛沛,曹扬,陈云明,等.陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征[J].生态学报,2017,37(2):443-454.
[23]屈凡柱,孟灵,付战勇,等.不同生境条件下滨海芦苇湿地C、N、P化学计量特征[J].生态学报,2018,38(5):1731-1738.
[24]陈文,王桔红,彭玉姣,等.不同生境中鬼针草(Bidenspilosa)碳氮磷化学计量特征及其营养利用策略[J].广西植物,2018,38(3):281-288.
[25]刘瑜,王庆贵,邢亚娟.温带森林叶部生态化学计量学特征研究进展[J].中国农学通报,2018,34(10):95-100.
[26]雍艳华.新疆四种藜科典型盐生植物及其生境土壤生态化学计量特征的比较分析[D].新疆:石河子大学,2017:1-73.
[27]阿布里孜·阿不都热合曼,吕光辉,张雪妮,等.新疆艾比湖流域植物光合器官碳、氮、磷生态化学计量特征[J].生态学杂志,2015,34(8):2123-2130.
[28]ELSERJJ,FAGANWF,KERKHOFFAJ,etal.Biologicalstoichiometryofplantproduction:metabolism,scalingandecologicalresponse toglobal change.[J]. NewPhytologist, 2010, 186(3):593-608.
[29]STERNERRW,ELSERJJ.EcologicalStoichiometry:TheBiologyofElementsfromMoleculestotheBiosphere.Princeton:PrincetonUniversityPress,2002.
[30] KOERSELMAN W, MEULEMAN A F M. The vegetation N∶P ratio: A new tool to detect the nature of nutrient limitation [J]. J ApplEcol, 1996, 33(6):1441-1450.
文章摘自:刘若璇,崔东,赵阳,刘海军,刘文新,张静。伊犁河谷不同生境下大麻根茎叶生态化学计量特征[J]环境化学,2022,41(5):1639-1648.
