摘 要:土壤污染问题是当前亟待解决的最严重的问题之一,在众多修复重金属污染土壤的方法中,植物修复技术是当前的热点,既通过植物修复技术对重金属污染的土壤进行修复。亚麻因生物量大、适应性强、对重金属具有较强的耐受性等,被认为是修复重金属土壤的理想材料。有关亚麻与重金属的研究,大多都是亚麻在重金属胁迫下的种子、幼苗的生长发育,转录组数据分析以及亚麻修复重金属污染土壤的研究,本综述主要是对以下四个方面进行综述总结:(1)重金属污染对植物生长的影响;(2)重金属污染物对亚麻生长的影响;(3)重金属污染对亚麻分子机制的影响;(4)重金属毒害亚麻的缓解措施。本综述通过对以上内容进行总结,为后期研究亚麻修复重金属污染土壤提供一定的理论依据。
关键词:亚麻;重金属污染;植物修复
土壤重金属污染是农业生产中亟待解决的重大难题。重金属污染土壤具有隐蔽性、不可逆转、长期性等特点(范占煌和董国云, 2015, 资源节约与环保, 2: 167-169),由于重金属较长时间停留在土壤中,导致土壤肥力下降,影响植物的生长发育,引起产量和品质下降,甚至污染地下水等(王秋平等, 2020; 陈奕暄等,2022)。常见的污染土壤的重金属元素主要有铅、镉、汞、铜、铬等金属元素(王元昌等, 2021),以上重金属元素具有较强的毒性和移动性,极易被吸收进入植物体内,阻碍植物的生长和发育(赵信林等, 2020),重金属离子通过降低蒸腾速率、气孔导度等影响光合作用,通过抑制叶绿素的生物合成、脂质过氧化等影响生物量(王秋平等, 2020),以及进入食物链进行富集,对动物和人体健康造成威胁。
对重金属污染的土壤进行修复,有四种修复类型,分别是工程修复、物理修复、化学修复和生物修复(王玉富等, 2015; 向魏, 2020),现目前修复重金属污染土壤的热点是生物修复。生物修复是以植物修复为主,植物修复属于一种原位、环保、低成本的修复技术(赵信林等, 2020; 杨耀东, 2021),具有低成本、高效益、来源广等优点(安霞等, 2019),被认为是修复重金属污染土壤最有效的方法(陈奕暄等, 2022)。植物修复是由植物根系通过选择性的吸收、运转和自身的生物富集、储存和降解,将土壤中的重金属固定于根部(范占煌和董国云, 2015, 资源节约与环保, 2: 167-169),就是利用超富集重金属的植物从污染土壤中去除重金属污染物(王玉富等, 2015; 柳婷婷等, 2022),超富集植物对重金属污染土壤具有较强的吸收能力和耐受能力,是重金属污染土壤的理想修复材料(王玉富等, 2015)。然而,较多的超富集植物因其生物量小,适应环境能力弱等特点(王玉富等, 2015; 柳婷婷等, 2022),严重限制了它们在修复领域的应用(杨耀东, 2021)。目前,用于植物修复的作物都是生物量大、适应性强、具有一定的经济价值的作物,利用经济作物修复重金属污染土壤是一种边利用边修复的理想方法(王元昌等, 2021)。亚麻具有较强的富集和转运重金属的能力,适合替代那些生长缓慢、生物量小的超积累植物,实现对重金属污染土壤的边修复边利用(陈玉梅, 2020; 杨耀东, 2021; 赵信林等, 2022)。
亚麻(Linum usttatissimum L.)是亚麻科亚麻属、一年生或多年生双子叶草本植物(曲志华等,2019;陆美光等,2021),是一种综合利用价值极高的广泛种植的经济作物,具有悠久的栽培历史(巩文靓等,2020;黄文功等,2021)。亚麻是世界五大油料作物和三大纤维作物之一,按其用途可分为油用亚麻、纤用亚麻和油纤 兼用亚麻(安霞等,2019;赵信林等,2022)。油用亚麻亦称为胡麻(党照等,2018),亚麻籽中有较高的含油量,亚麻油是一种保健食用油,富含多种对人体有用的营养成分,亚麻油中含有α-亚麻酸,是人体必需的一种不饱和脂肪酸,具有较高的经济价值和营养价值(赵东晓等,2020;柳婷婷等,2022)。亚麻籽中含有丰富的木酚素、果胶、亚麻胶等营养成分,被广泛用于食品、化妆品和保健品等领域(赵东晓等,2020;李世玉等,2021)。纤用亚麻具有强韧性、吸湿、透气等特点(巩文靓等,2020),可用于造纸和纺织等行业,具有“纤维皇后”之称(伍叶娜等,2022)。因纤用亚麻具有生物量高,适应性强,不进入食物链传递,对重金属污染土壤具有较强的耐受性等特点(柳婷婷等,2022),被认为是一种修复重金属污染土壤的理想作物(巩文靓等,2020;王元昌等,2021)。在南方可冬季种植亚麻,与其他作物进行轮作,有效提高冬闲田的经济价值,是一种理想的边利用边修复的作物(王玉富等,2015;柳婷婷等,2022),亚麻是重金属镉污染农田中适合种植的替代经济作物(柳婷婷,2022)。
1 重金属污染对植物生长的影响
重金属胁迫不仅抑制植物地上部的生长,还抑制地下部的生长(Park et al.,2015)。重金属胁迫对植物生长的影响,主要是对种子萌发、幼苗生长及生理生化指标等产生影响,重金属胁迫时,植物会产生大量的自由基破坏细胞膜,破坏保护酶系统,影响植物的抗性(葛坤等,2021);植物通过产生生物效应,使蛋白质变形、生物酶活性降低,阻碍植物对必须营养元素的吸收,导致植物体内的生理代谢紊乱,产生大量的活性氧自由基,抑制植物生长,严重时导致植物死亡(范占煌和董国云,2015,资源节约与环保,2:167-169)。重金属胁迫时,重金属通过干扰植物对养分的吸收和分配,导致植物的养分缺乏或失衡等,产生氧化应激反应,最终影响植物的生长(王秋平等,2020)。过量的重金属对植物生长造成严重的影响,植物出现叶片失绿、坏死、膨压丧失、腐烂、种子发芽率降低、光合器官受损、甚至植物死亡等现象(Park et al.,2015)。
重金属对植物的胁迫主要是植物体内抗氧化酶活性的变化,适量的重金属可提高抗氧化酶活性,超过临界值则对植物生长不利,甚至导致植物死亡(吴嘉煜和米楠,2022)。在铜胁迫时,直接影响植物的光合作用、叶绿素含量、气孔运动、抗氧化剂的产生和酶活性等(向魏,2020),从而抑制植物幼苗的生长,降低芽长、根长及幼苗鲜重(向魏等,2022)。锌胁迫对植物产生的影响是:根和茎生物量下降,叶绿素降解,嫩叶卷曲和叶尖死亡等症状,当高浓度时,光合参数会发生变化,显著抑制植物的生长(Saraswat and Rai,2011)。锌胁迫时,当锌处于适当浓度时能提高植物种子的代谢水平,利于植物的生长发育;当浓度过高会加速产生大量的活性氧,对种子储存营养物质造成氧化损伤,影响种子萌发和幼苗生长(李世玉等,2021)。镉胁迫会导致植物生长迟缓、植株矮小、缺绿、产量下降(柳婷婷,2022),镉胁迫对植物生理的影响主要是:改变酶活性和结构、脂质过氧化、降低光合作用、黄化、氮代谢、蒸腾作用、种子萌发、根损伤、以及植物死亡等(Saraswat and Rai,2011);镉胁迫还会干扰植物的光合作用、气体交换、养分吸收等生理活动,导致植物生长和生物产量下降(Singh et al.,2016)。镉对种子萌发的影响,主要是通过抑制种子中淀粉酶活性,降低可溶性糖和还原糖含量,减少所需的物质和能量,影响发芽率及后期幼苗的生长(熊敏先等,2021)。
2 重金属污染对亚麻生长的影响
2.1 重金属污染对亚麻生长的影响
正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容亚麻品种对重金属的反应具有高度的多样性,不同品种的毒性差异较大(Soudek et al.,2010)。研究表明:对重金属镉的吸收和富集能力,亚麻强于棉花、大麻,在镉污染土壤中亚麻可作为适合种植的替代经济作物(柳婷婷等,2022)。亚麻对重金属具有较强的耐性,能较好的吸收,并在植物体内进行积累,实现亚麻对土壤重金属的富集(王玉富等,2015)。亚麻对镉的富集系数大于1,根和茎基部中镉含量最高,茎的富集系数为13.3(Douchiche et al.,2012;王玉富等,2015)。亚麻对重金属污染的土壤具有较高的耐受性,低浓度时不会影响亚麻的生长,不同品种对重金属的耐受性不同(柳婷婷等,2022),亚麻品种与重金属之间具有互作性(王玉富等,2015)。不同重金属离子在亚麻的毒性效应不同(安霞等,2020),不同重金属在亚麻中的最高含量不同,镉主要是位于根中,锌主要是位于茎中,铅和铜主要是位于蒴果中(王玉富等,2015)。不同重金属在亚麻植株内的转移能力不同,同种重金属在不同亚麻品种中的转移能力也不同(王玉富等,2015)。
2.1.1镉胁迫对亚麻生长的影响
重金属镉是农田土壤中主要的污染元素之一,镉是一种毒性大、移动性强、易被植物吸收的重金属元素(柳婷婷等,2022;赵信林等,2022)。赵信林等(2022)研究发现,镉处理会影响亚麻的出苗率、株高、茎粗、叶片数,最终影响生物量。镉胁迫会抑制农作物的生长,导致作物出现生长迟缓、植株矮小、缺绿、产量下降等现象(孙丽娟等,2018)。不同浓度的镉胁迫会影响亚麻种子的发芽过程(安霞等,2020)。镉胁迫对亚麻的影响,可通过不同的镉浓度处理影响亚麻种子的发芽率,低浓度(<100mg/L)时,对发芽率影响不大,但当镉浓度升高后(>100mg/L),就会降低亚麻种子的发芽率,镉胁迫浓度越高对亚麻种子发芽率的降低作用越明显(安建平等,2008)。镉处于低质量分数时促进亚麻的出苗,当处于高分子量时,对亚麻的出苗率起抑制作用(赵信林等,2022)。镉处理对亚麻的株高、茎粗、叶片数和生物量起抑制作用,抑制作用与镉浓度成正相关(赵信林等,2022)。镉胁迫时,亚麻各器官中镉含量为:根>叶>茎,细胞壁、核糖体中的含量显著高于细胞核、叶绿体及线粒体细胞中的镉含量(赵信林等,2022)。
镉胁迫会抑制亚麻幼苗的生长,导致亚麻幼苗的组织增大,体积保持不变,细胞壁增厚和下表皮折叠引起细胞改变(Douchiche et al., 2007;王玉富等,2015)。当镉处于低浓度时,刺激亚麻种子内RNA和蛋白质的合成,利于种子萌发和出苗(赵信林等,2022)。当处于高浓度时,会严重降低亚麻的生物量,降低速率与镉浓度呈正比(赵信林等,2022),导致该现象可能是活性氧产生导致酶和官能团堵塞或失活,抑制植物光合作用,导致生物量减少(赵信林等, 2022)。镉胁迫时,会抑制亚麻对钾、钙、镁、铁的吸收,增加亚麻的根部和地上部对镉的积累(Belkhadi et al.,2010)。当镉胁迫达到一定浓度时,植物就会产生大量的活性氧,提高了抗氧化酶系统的清除能力(张星雨等,2021),早期清除活性氧是植物耐镉性的重要组成部分(Tamás et al.,2015)。镉浓度对亚麻脂肪酶的影响,低浓度时提高亚麻脂肪酶的活性,高浓度时则会降低脂肪酶活性(安建平等,2008)。亚麻植株具有较强的耐镉性,表现在亚麻根部限制镉向上运输、以及细胞壁和液泡对镉的隔离等(赵信林等,2022)。镉胁迫时,亚麻植株中的丙二醛含量增加,叶片会出现较高的脂质过氧化(Belkhadi et al.,2010)。在较高镉浓度时,活性氧具有较高活性和毒性,对膜脂质、蛋白质、核酸、以及叶绿体、线粒体、细胞膜等会造成损伤,导致植物死亡;活性氧还可通过改变膜流动性和渗透性等,从而降低细胞活性(Singh et al.,2016)。镉胁迫时,会改变亚麻植株中的不饱和脂肪酸组成,降低亚麻的脂肪酸总量(Park et al.,2015)。
2.1.2其他重金属胁迫对亚麻生长的影响
铜胁迫对亚麻种子活力的影响,亚麻种子活力与铜胁迫浓度呈反比,铜对亚麻萌发有抑制作用,低浓度时可显著促进亚麻生长,但当铜浓度达到一定浓度时(≥50 mg/L)会抑制萌发,降低亚麻的发芽率(向魏等,2022);铜溶液会显著抑制亚麻主根的生长,铜胁迫浓度与亚麻主根抑制指数、芽长抑制指数均呈正比关系(向魏等,2022)。高浓度的铜,通过降低植株高度、直径、鲜干生物量和叶片叶绿素含量等途径影响植物的生长发育,显著影响了植物的生长和生物量,亚麻能够从土壤中除去大量的铜(Saleem et al.,2020)。
铬胁迫对亚麻生长的影响,王玉富等(2015)综合总结:铬主要是影响亚麻植株的分枝数,当铬为低浓度时(25mg/kg),对亚麻的分枝数影响不显著,在一定程度上有利于亚麻的生长,可促进亚麻植株的株高、叶绿素浓度等;当铬为高浓度时(>125mg/kg),会显著降低亚麻植株的分枝数,蒴果数随铬浓度的升高而显著降低(王玉富等,2015)。
铅对亚麻生长的不利影响有降低亚麻植株的株高、抑制生物量,对铅的积累量的根部>地上部,随铅浓度胁迫的升高,超氧化物歧化酶和过氧化物酶活性先升高后降低的变化趋势(Pan et al.,2020)。低钾处理时,亚麻叶片边缘变黄、植株矮化(黄文功等,2021)。
2.2重金属在亚麻体内的分布
亚麻的根和叶中的镉含量较高,叶和茎中的镉含量随时间呈减少的趋势(House et al.,2020)。重金属吸附的主要器官是亚麻根系(王元昌等,2021),亚麻植株体内重金属的积累浓度为:根>茎>蒴果>种子,富集能力为:根>茎>叶>籽(王玉富等,2015)。重金属在作物轴上的分布具有选择性,亚麻中的中重金属含量为:根>茎>叶>种子(Angelova,2004)。
镉在亚麻四个器官中的平均积累量为:韧皮部>根>木质部>蒴果,镉积累的主要部位是亚麻的韧皮部,不同亚麻品种的镉吸收量与生物量之间呈极显著正相关(Guo et al.,2020)。不同用途的亚麻品种对镉的富集能力不同,纤用亚麻与油用亚麻品种相比,纤用亚麻具有更强的重金属镉富集能力(王玉富等,2015)。镉胁迫下,亚麻植株中镉的含量是地上茎叶<地下根(王玉富等,2015),富积系数大于1,对镉具有一定的富集能力(许英等,2007),随土壤中重金属镉浓度的增加,亚麻植株体内的镉含量也会显著增加(赵信林等,2022)。亚麻对重金属镉具有较高的耐受性和富集作用,亚麻叶中的转运系数最高,对镉的富集能力最强,亚麻具有较强的从根部往地上部转运重金属镉的能力(陈奕暄等,2022)。
3 重金属胁迫对亚麻酶系统的影响
适度的重金属胁迫能提高植物的抗氧化酶活性,当浓度过高则会影响活性,严重时导致植物死亡(吴嘉煜和米楠,2022)。植物的抗氧化酶系统主要包括:过氧化氢酶、过氧化物酶和超氧化物歧化酶等,抗氧化酶活性与植物的抗逆性呈正相关(吴嘉煜和米楠,2022)。镉离子污染会使植物叶片的超氧化物歧化酶、过氧化氢酶、过氧化物酶和硝酸还原酶的活性下降,低镉浓度能提高亚麻萌发的脂肪酶活性,增强脂质代谢的能力,高镉浓度时能抑制亚麻的代谢(安建平等,2008)。当镉积累到一定程度时植物会产生大量的活性氧,抗氧化酶系统中的酶就负责清除细胞内的活性氧(张星雨等,2021),活性氧大量累积使各种酶和官能团堵塞或失活,抑制植物的光合作用(赵信林等,2022)。镉胁迫导致亚麻幼苗的过氧化氢酶活性下降,因此该酶活性下降可能与H2O2含量增加有关(徐严等,2012)。铅胁迫时,亚麻幼苗中的脯氨酸、丙二醛含量升高,随处理时间的增加含量越高,亚麻植株中的过氧化氢酶活性、过氧化物酶活性、超氧化物歧化酶的活性先升高后逐渐降低,会导致细胞受到氧化损伤(李宇洁和安梅,2013)。铜胁迫时,会引起活性氧物质的产生,亚麻根和叶中的超氧化歧化酶和过氧化物酶活性增加(Saleem et al.,2020)。
4 第四章
亚麻在重金属污染土壤中仍可正常生长,因其具有较强的耐受性,在重金属胁迫时会产生应激机制,降低镉产生的影响(王玉富等,2015)。在分子水平上,植物主要是通过胁迫相关蛋白和信号分子的合成来响应镉胁迫图1 (Singh et al.,2016),参与植物镉胁迫响应的主要有五种,分别是:锌/铁转运蛋白、重金属ATP酶、ATP结合盒转运蛋白、阳离子扩散促进因子和天然抗性噬菌细胞蛋白等(张星雨等,2021)。亚麻耐镉机制是铁蛋白和小分子的巯基肽与镉结合,减少镉胁迫对亚麻的危害(王玉富等,2015)。镉胁迫时,植物体内的钙离子浓度升高,刺激钙调蛋白的表达,可促进离子转运、基因调节、代谢和应激耐受性等(Singh et al.2016)。
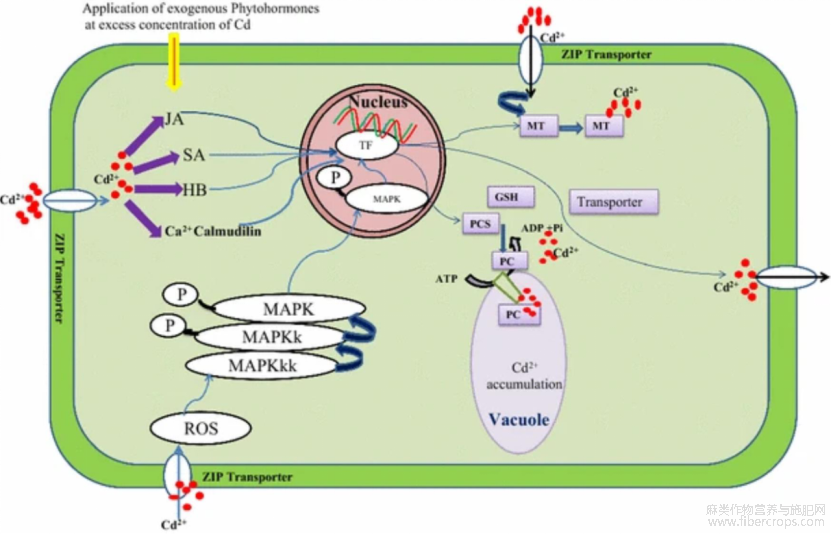
图1内源和外源植物激素作用下镉信号转导图示(Singh et al., 2016)
Figure 1 Cadmium signal transduction diagram under the action of endogenous and exogenous plant hormones (Singh et al., 2016)
注:JA: 茉莉酸; SA: 水杨酸; BR: 油菜素类固醇; TF: 转录因子; MT: 金属硫蛋白; PCS: 植物螯合素; GSH: 谷胱甘肽; ROS: 活性氧; MAPK: 丝裂原活化蛋白激酶; MAPKKK: MAPK激酶激酶; MAPKK: MAPK激酶Note: JA: Jasmonic acid ; SA: Salicylic acid; BR: Brassinosteroid; TF: Transcription factor; MT: Metallothionein; PCS: phytochelatin; GSH: Glutathione; ROS: Active oxygen; MAPK: Mitogen-activated protein kinase; MAPKKK: MAPK kinase kinase, MAPKK: MAPK kinase
植物通过上调果胶甲酯酶(PME)基因增加对重金属镉的适应性,通过上调SlPME1基因的表达,降低果胶甲酯化程度,减少镉进入原生质,加强植物对重金属镉胁迫的抵抗(张星雨等,2021)。镉胁迫时,植物根系可通过上调QUA1和PME1基因的表达,增加果胶和PME的合成,减少对细胞的破坏(张星雨等,2021)。CsHMA3的过量表达,是重金属胁迫下种子在数量和质量上比野生型更耐重金属,以及具有更好的脂肪酸组成的原因,该基因的过表达影响了植物中的脂肪酸组成和含量(Park et al., 2015)。在亚麻基因组中成功导入αMT1a基因(金属硫蛋白的α区域编码基因)并稳定表达,在镉污染土壤中,该转基因阳性亚麻株系对镉有较高的耐受能力(安霞等,2019)。
亚麻种子中镉积累的基因型差异,可能是部分镉在种子中的再分配差异(House et al.,2020)。亚麻ABC转运蛋白和HMA基因在ATP结合、转运、催化活性、ATP酶活性和金属离子结合等方面起着重要的作用(Khan et al.,2020)。亚麻响应重金属胁迫的方式,可能主要是通过信号转导途径,调节关键基因的表达量或对蛋白质进行修饰等,通过KEGG富集分析,发现亚麻在铅胁迫下会影响核苷酸的代谢途径(安霞等,2019)。亚麻品种对铅胁迫的耐受性与重金属胁迫相关基因的转录水平升高有关,对铅胁迫有较强的耐受性的四个标记基因分别为LuACBP、LuSOD1、LuPOD1和LuPOD2 (Pan et al.,2020)。在不同重金属胁迫,针对亚麻等多种作物的研究中,发现参与重金属胁迫响应和抗性过程的转录因子家族相关基因有TCP、GRF、NAC、MYB及BhLH等(安霞等,2020)。黄文功等(2021)研究亚麻响应低钾胁迫转录谱分析发现,3个低钾响应强烈的钾运转蛋白基因,分别是:LusKC1 (Lus K channel 1)、LusSKOR (Lus STELAR K+ outward rectifier)和LusHAK5 (Lus high affinity K+ transporter 5),当低钾处理12 h,共鉴定出1 154个差异表达基因,GO功能富集分析发现,差异基因富集于代谢过程、细胞进程、单一生物过程、催化活性和结合功能五大类,由KEGG通路富集分析,差异表达基因涉及的代谢通路有能量代谢、碳水化合物代谢、碳代谢、氨基酸代谢、萜类化合物代谢等通路(黄文功等,2021)。
5 重金属毒害亚麻的缓解措施
5.1 植物激素
植物激素可诱导植物应对重金属的毒害,如:生长素、细胞分裂素、水杨酸、茉莉酸和甾类激素等(Singh et al.,2016)。重金属胁迫时,植物通过将金属隔离在液泡内,改变根际的酸碱度、结合金属硫蛋白和刺激不同抗氧化酶等,从而减少植物对重金属的积累(Islam and Sandhi,2022)。根际微生物可通过植物激素调节、产生有机酸、氧化还原作用等机制,促进植物的生长增强其适应性,减轻重金属污染对植物的危害(王秋平等,2020)。根际促生菌产生的生长素可诱导植物的生理变化,提高植物对重金属污染的适应和耐受性,减轻重金属污染的毒害,另一方面,可通过促进植物根系增殖,增强植物对营养物质的吸收从而促进植物的生长(王秋平等,2020)。
植物体内的防御机制可以可控制金属的吸收、转运、积累和解毒,亚麻与菌根真菌可联合修复镉胁迫(Amna et al.,2015)。在重金属胁迫时,植物体内的乙烯含量增加,阻碍侧根的生长、根系伸长和根毛形成,抑制植物的生长(王秋平等,2020)。除此之外,根际促生菌可产生如水杨酸、赤霉素、细胞分裂素等植物激素,帮助植物适应重金属胁迫(王秋平等,2020)。水杨酸可通过幼苗,水杨酸可增加叶绿素、丙二醛和总脂的含量,可提高亚麻植株中的亚麻酸含量,可提高植物对镉胁迫的抗性(Belkhadi et al.,2010)。在镉胁迫时,使用乙酰水杨酸处理幼苗期的亚麻,可提高亚麻的光合作用,促进亚麻镉污染环境的适应(王玉富等,2015)。水杨酸会减低亚麻对镉的吸收和转运,增加亚麻对其他金属的吸收,有利于植物的生长(Belkhadi et al.,2010)。水杨酸预处理,对亚麻生长、镉积累和叶绿素含量有促进作用,对亚麻幼苗镉毒害具有保护作用(Belkhadi et al.,2010)。水杨酸通过抑制镉诱导的生长素介导活性氧产生,阻止或部分抑制根系对镉的防御反应(Tamás et al.,2015)。水杨酸通过调节亚麻幼苗的脂质代谢、抗氧化系统等,间接缓解镉的毒害,从而改善镉胁迫下植物的生长状况(Belkhadi et al.,2010)。
5.2 外源NO
NO可清除植物体内活性氧,促进物质的积累,调节抗氧化酶的活性(李宇洁和安梅,2013)。亚麻幼苗在镉胁迫下可通过使用外源NO处理,可显著降低幼苗的丙二醛含量,提高亚麻幼苗的超氧化物歧化酶、过氧化物酶和过氧化氢酶的活性,加快对活性氧的清除,对细胞膜、亚麻幼苗具有保护作用,可减轻镉胁迫对亚麻幼苗的伤害(徐严等,2012)。外源NO能促进铅胁迫下亚麻亚苗幼苗叶片中的丙二醛、脯氨酸含量积累,提高抗氧化酶系统的活性,缓解亚麻叶片的氧化损伤等,可增强亚麻抗重金属铅的能力(李宇洁和安梅,2013)。氮、锌处理均可提高亚麻种子产量,降低亚麻种子中镉的含量,提高亚麻品质(Amna et al.,2015)。
5.3 有机酸
正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容正文内容不同有机酸种类及用量都可以不同程度地增加亚麻的株高和单株重的趋势。根际促生菌对外部不利环境有一定的抗性,可增强植物对重金属污染的修复潜力(石杨等,2022),可产生草酸、乙酸和柠檬酸等有机酸,通过形成螯合物帮助重金属污染土壤中的植物生长,减轻重金属的危害(王秋平等,2020)。赵信林等(2020)研究发现有机酸为0.6mmol时,抑制土壤中重金属镉的活性,亚麻不受镉的毒害;当有机酸为1.2mmol时,镉的活性增强,亚麻生长受到抑制;当有机酸为2.4 mmol时,镉活性增强,亚麻对镉的耐性提高,在一定程度上可提高亚麻的株高(赵信林等,2020)。柠檬酸、苹果酸和草酸对亚麻株高的影响比对茎粗和单株重的影响更显著,高浓度苹果酸和草酸处理可提高亚麻植株的吸镉量,亚麻对土壤中Cd的移除效果增强,低浓度的草酸可提高亚麻对镉的转移系数和富集系数(赵信林等,2020)。通过控制根部金属的吸收和远距离金属运输等方式,从而实现植物对重金属毒害的保护(Belkhadi et al.,2010)。
5.4 田间管理
采取科学施肥、合理利用有机肥和适当的种植结构调整等,可缓解和消除重金属对植物造成的危害(吴嘉煜和米楠,2022)。亚麻对重金属具有较强的耐受性和吸附作用,通过适当的调节剂、水分、肥料、微生物等的使用和调节,可提高亚麻对重金属的吸收、转运和吸附能力(王玉富等,2015)。在轮作修复重金属污染土壤中,可采用合适的耕作模式、使用适当化肥、水分管理以及杂草清理等可增加产量提高重金属的提取量(陈玉梅,2020)。在重金属镉污染的土壤中,丛枝菌根真菌能创造出适合亚麻根系生长的环境,促进根系的生长,可消除耕地中重金属对种子产量的影响(王玉富等,2015)。根际促生菌可维持土壤肥力、促进植物生长,对重金属胁迫有一定的抗性(石杨等,2022),通过植物激素、螯合作用、提高对氧化还原和合成抗氧化酶等,缓解重金属胁迫下植物的应激机理,缓解重金属的毒害影响(王秋平,2020)。增加重金属污染土壤的湿度,提高了土壤中镉的流动性,增加亚麻植株对镉的积累,减少土壤中镉的含量(王玉富等,2015)。施用磷肥可增加亚麻对重金属镉的吸收,通过作物轮作与磷肥管理等方式降低重金属污染对亚麻植株的影响,种植顺序和磷素管理可以通过提高Zn含量、降低Cd含量来改善亚麻籽的食品品质(Grant et al.,2010;王玉富等,2015),施加磷肥降低了作物体内锌的积累,增加了镉的吸收和转运,从而间接增加作物体内镉的含量,耕作制度、前茬作物和磷肥可能通过影响菌根定殖来影响Cd和Zn的积累(Grant et al.,2010)。有机物料的使用能改变土壤微环境影响重金属的存在形态,可缓解重金属胁迫,促进植物生长、增强植物的抗逆性(吴嘉煜和米楠,2022)。
6 结语
在重金属胁迫下,亚麻的生长、生理以及分子方面都受到了不同程度的影响。重金属处于低浓度时,对亚麻的生长影响不大,当达到一定浓度时会严重影响亚麻植株的生长,甚至导致植株死亡。重金属污染对亚麻产生的毒害,可通过植物激素、外源NO、有机酸、以及田间管理等措施来缓解毒害情况。亚麻在重金属污染的土壤中可将重金属富集或转移在植株体内,减少或清除重金属的污染,达到修复修复重金属污染土壤的效果。因此,对重金属污染土壤中的亚麻,实行规模化、机械化种植,利于集中处理亚麻原料等(王玉富等,2015),可加强亚麻修复重金属污染土壤。加大引进高富集的亚麻品种,筛选用于修复重金属污染土壤中的亚麻品种及种质资源,以及通过结合基因工程选育高富集的亚麻品种(王玉富等,2015),结合分子生物学理念寻找耐重金属的基因和转运蛋白,通过转基因技术在植物中提高植物对重金属污染的修复效果(孙丽娟等,2018;时唯伟,2022);其次充分利用动物、植物、微生物联合修复重金属污染的土壤(Mukhtar et al.,2019),通过寻找良种良法配套使用,达到亚麻对重金属污染土壤的最佳修复效果。本综述可为亚麻修复重金属污染土壤的研究提供一定的基础。
作者贡献
刘琴是综述的主要撰写人,完成相关文献资料的收集和分析及论文初稿的写作;安霞、杜光辉参与文献资料的分析、整理;安霞是项目的构思者及负责人,指导论文写作。全体作者都阅读并同意最终的文本。
致谢
本研究由国家重点研发计划政府间国际科技创新合作重点专项(2017YFE0195300)、国家自然科学基金(31801406, 31872130, 32202506)、浙江省基础公益研究计划(LGN20C150007)、财政部和农业农村部:国家麻类现代农业产业技术体系(CARS-16-S05)和浙江省农业科学院国际/港澳台合作专项经费(2022)共同资助。
参考文献
[1] Amna., Ali N., Masood S., Mukhtar T., Kamran M.A., Rafique M., Mussain M.F.H., and Chaudhary H.J., 2015, Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake ofLinum usitatissimum in association with Glomus intraradices. Environ Monit Assess, 2015, 187: 1-11.
[2] [An J.P., Wang T.P., Xie T.Z., Yang A.M., and Wang L., 2008, Effect of Cd stress on seed germination and seedling physiological characteristics of flax(Linum usitatissimum L.), Zhongguo Maye Kexue (Plant Fiber Sciences in China), 30(4): 199-203. (安建平, 王廷璞, 谢天柱, 杨爱民, 王莉, 2008, 镉胁迫对亚麻种子萌发及幼苗生理特性的影响, 中国麻业科学, 30(4): 199-203.)
[3] An X., Jin G.R., Chen J., Luo X.H., Chen C.L., Li W.L., and Zhu G.L., 2020, Transcriptome characterization analysis ofLinum usitatissimum under Pb stress, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 18(10): 3185-3191. (安霞, 金关荣, 陈杰, 骆霞虹,陈常理, 李文略, 朱关林, 2020, 亚麻(Linum usitatissimum)铅胁迫转录组初步分析, 分子植物育种, 18(10): 3185-3191.)
[4] An X., Jin G.R., Li L.F., Luo X.H., Chen C.L., Li W.L., Zhu G.L., and Zhang J.Y., 2019, Research progress on genetic engineering of flax (Linum usitatissimum L.), Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 17(20): 6705-6710. (安霞, 金关荣, 李鲁峰, 骆霞虹, 陈常理, 李文略, 朱关林, 张景瑜, 2019, 亚麻基因工程研究进展, 分子植物育种, 17(20): 6705-6710.)
[5] Angelova V., Ivanova R., Delibaltova V., and Ivanov K., 2004, Bio-accumulation and distribution of heavy metals in fibre crops (flax, cotton and hemp), Ind Crop Prod, 19(3): 197-205.
[6] Belkhadi A, Hediji H, Abbes Z, Nouairi I., Barhoumi Z., Zarrouk M., Chaibi W., and Djebali W., 2010, Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content inLinum usitatissimum L., Ecotox. Environ. Safe, 73(5): 1004-1011.
[7] Chen Y.M., 2020, Remediation of low cadmium-polluted cultivated land by several crop rotation systems, Thesis for M.S., College of Resources and Environment, Hunan Agricultural University, Supervisor: Zeng Q.R., pp.13-18.(陈玉梅, 2020, 几种作物轮作模式对镉轻度污染耕地修复研究, 硕士学位论文, 湖南农业大学资源植物环境学院, 导师: 曾清如, pp.13-18.)
[8] Chen Y.X., Deng X., Yang Y., and Zeng Q.R., 2022, The potential of flax-rice rotation for remediation of cadmium contaminated soil, Zuowu Yanjiu (Crop Research), 36(2): 126-131. (陈奕暄, 邓潇, 杨洋, 曾清如, 2022, 亚麻—水稻轮作模式对镉污染土壤修复潜力研究, 作物研究, 36(2): 126-131.)
[9] Dang Z., Zhang J.P., Wang L.M., Zhao W., and Li W.J., 2018, Correlation between environmental factors and yield and quality of flax, Gansu Nongye Keji (Gansu Agricultural Science and Technology), (6): 21-25. (党照, 张建平, 王利民, 赵玮, 李闻娟, 2018, 环境因子与胡麻产量及品质相关性研究, 甘肃农业科技, 6: 21-25.)
[10] Douchiche O., Chaïbi W., and Morvan C., 2012, Cadmium tolerance and accumulation characteristics of mature flax, cv. Hermes: Contribution of the basal stem compared to the root, Journal of Hazardous Materials, 235: 101-107.
[11] Douchiche O., Rihouey C., Schaumann A., Driouich A., and Morvan C., 2007, Cadmium-induced alterations of the structural features of pectins in flax hypocotyl, Planta, 225: 1301-1312.
[12] Ge K., Wang P.J., Shao H.L., Guo J.Y., and Du B., 2021, Study on the effects of heavy metal stress on plant physiology and biochemistry and its resistance mechanism, Shanxi Linye Keji (Shanxi Forestry Science and Technology), 50(3): 43-46. (葛坤, 王培军, 邵海林, 郭家雁, 杜宾, 2021, 重金属胁迫对植物生理生化的影响及其抗性机理研究, 山西林业科技, 50(3): 43-46.)
[13] Gong W.L., Kang X.M., Ma M.Y., Duan H.Y., and Jiang G.H., 2020, Research progress on QTL mapping of flax, Zhongguo Maye (Plant Fiber Sciences in China), 42(4): 187-192. (巩文靓, 康雪蒙, 马梦影, 段海燕, 姜恭好, 2020, 亚麻QTL定位的研究进展, 中国麻业科学, 42(4): 187-192.)
[14] Grant C.A., Monreal M.A., Irvine R.B., Mohr R.M., Mclaren D.L., and Khakbazan M., 2010, Preceding crop and phosphorus fertilization affect cadmium and zinc concentration of flaxseed under conventional and reduced tillage, Plant and Soil, 333(1-2): 337-350.
[15] Guo Y., Qiu C.S., Long S.H., Wang H., and Wang Y.F., 2020, Cadmium accumulation, translocation, and assessment of eighteenLinum usitatissimum L.cultivars growing in heavy metal contaminated soil, Int. J. Phytoremediat, 22(5): 490-496.
[16] [16] House M.A., Young L.W., Liu X., Liber K., Diederichsen A., and Booker H.M., 2020, Comparative analysis of cadmium uptake and distribution in contrasting canadian flax cultivars, BMC Res Notes, 13(1): 1-6.
[17] Huang W.G., Jiang W.D., Yao Y.B., Song X.X., Liu Y., Chen S., Zhao D.S.,Wu G.W., Yuan H.M., Ren C.Y., Sun Z.Y., Wu J.Z., and Kang Q.H., 2021, Transcriptiome profiling of flax (Linum usttatissimum L.) response to low potassium stress, Zuowu Xuebao (Journal of Crops), 47(6): 1070-1081.(黄文功, 姜卫东, 姚玉波, 宋喜霞, 刘岩, 陈思, 赵东升, 吴广文, 袁红梅, 任传英, 孙中义, 吴建忠, 康庆华, 2021, 亚麻响应低钾胁迫转录谱分析, 作物学报, 47(6): 1070-1081.)
[18] Islam M., and Sandhi A., 2022, Heavy metal and drought stress in plants: The role of microbes-A review, Gesunde. Pflanzen., 2022: 1-14.
[19] Khan N., You F.M., Datla R., Ravichandran S., Jia B., and Cloutier S., 2020, Genome-wide identification of ATP binding cassette (ABC) transporter and heavy metal associated (HMA) gene families in flax (Linum usitatissimum L.), BMC genomics, 21: 1-14.
[20] Li S.Y., Lu X.P., Dai Z.G., Wang H.F., Su J.G., Sun J., and Xie D.W., 2021, Effects of Exogenous Zinc on Nutrition and Harmful Components of Flax Sprout, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 43(4): 190-197. (李世玉, 陆旭鹏, 戴志刚, 王华峰, 粟建光, 孙健, 谢冬微, 2021, 外源锌对亚麻芽菜营养及有害成分的影响, 中国麻业科学, 43(4): 190-197.)
[21] Li Y.J., and An M., 2013, Protective effect of exogenous nitric oxide on sseeding of linseed under Pb stress, Gansu Nongye Keji (Gansu Agricultural Science and Technology), 2013(11): 26-29. (李宇洁, 安梅, 2013, 外源NO对Pb胁迫下亚麻幼苗的保护效应, 甘肃农业科技, 2013(11): 26-29.)
[22] Liu T.T., Luo X.H., Li W.L., Zhu G.L., and An X., 2022, Difference analysis of Cd absorption and accumulation in different flax varieties, Zhejiang Nongye KeXue (Zhejiang Agricultural Sciences), 63(9): 2030-2032+2045.(柳婷婷,骆霞虹,李文略,朱关林,安霞, 2022, 不同亚麻品种重金属Cd吸收积累的差异, 浙江农业科学, 63(9): 2030-2032+2045.)
[23] Lu M.G., Duan H.Y., and Jiang G.H., 2021, Flax cenome association analysis: a review, Zhongguo Nongxue Tongbao (Chinese Agricultural Science Bulletin), 37(21): 111-118.(陆美光, 段海燕, 姜恭好, 2021, 亚麻全基因组关联分析的研究进展, 中国农学通报, 37(21): 111-118.)
[24] Mukhtar Y., Galalain A.M., Abdu K., Tukur S., Bashir R.A., Abdulkadir A.I., and Habib A.A., 2019, A comprehensive review on phytoremediation potentials ofHibiscus cannabinus Linn. (kenaf): mechanisms, benefits and prospects, American Journal of Natural Sciences, 2(1): 10-23.
[25] Pan, G., Zhao, L.N., Li J.J., Huang S.Q., Tang H.J., Chang L., Dai Z.G., Chen A.G., Li D.F., Li Z., and Deng Y., 2020, Physiological responses and tolerance of flax (Linum usitatissimum L.) to lead stress, Acta. Physiol. Plant, 42:113.
[26] Park W., Feng, Y.f., Kim H., Suh M.C., and Ahn S.j., 2015, Changes in fatty acid content and composition between wild type and CsHMA3 overexpressingCamelina sativa under heavy-metal stress, Plant Cell Rep, 34: 1489–1498.
[27] Qu Z.H., Bai W., Zhang L.L., Li F., Hu Y., and Qiao H.M., 2019, Main agronomic characteristics analysis on 170 flax germplasm resources, Zuowu Zazhi (Crops), (4): 77-83. (曲志华, 白苇, 张丽丽, 李峰, 胡杨, 乔海明, 2019, 170份亚麻种质资源主要农艺性状分析, 作物杂志, (4): 77-83.)
[28] Saleem M.H., Fahad S., Khan S.U., Din M., Ullah A., Sabagh A.E., Hossain A., Llanes A., and Liu L., 2020, Copper-induced oxidative stress, initiation of antioxidants and phytoremediation potential of flax (Linum usitatissimum L.) seedlings grown under the mixing of two different soils of China, Environ Sci Pollut Res, 27: 5211-5221.
[29] Saraswat S., and Rai J.P.N., 2011, Complexation and detoxification of Zn and Cd in metal accumulating plants, Rev Environ Sci Biotechnol, 10: 327-339.
[30] Shi W.W., Zhou C.X., Liu K., and Wang R., 2022, Study on phytoremediation of heavy metal Cd-contaminated soil, Zhongguo Ziyuan Zonghe Liyong (China Resources Comprehensive Utilization), 40(9): 93-95. (时唯伟, 周长行, 刘凯, 王榕, 2022, 重金属Cd污染土壤的植物修复研究, 中国资源综合利用, 40(9): 93-95.)
[31] Shi Y., Li J.H., Yu Y., Yang Y.W., Li B., Chen S.Q., Chen J., Zhao K., and Huang J., 2022, Progress and prospect of plant remediation technology joint with other technologies for heavy metal contaminated soil, Huanjing Wuran yu Fangzhi (Journal of Environmental Pollution and Prevention), 44(2):244-250. (石杨,李家豪,于月, 杨雨薇, 李斌, 陈思奇, 陈稷, 赵珂, 黄进, 2022, 重金属污染土壤的植物修复技术与其他技术联用的进展与前景, 环境污染与防治, 44(2): 244-250.)
[32] Singh S., Singh A., Bashri G., and Prasad S.M., 2016, Impact of Cd stress on cellular functioning and its amelioration by phytohormones: An overview on regulatory network, Plant Growth Regul., 80: 253-263.
[33] Soudek P., Katrusáková A., Sedlácek L., Petrová S., Kocí V., Marsík P., Griga M., and Vanek T., 2010, Effect of heavy metals on inhibition of root elongation in 23 cultivars of flax (Linum usitatissimum L.), Archives of Environmental Contamination and Toxicology, 59: 194-203.
[34] Sun L.J., Qin Q., Song K., Qiao H.X., and Xue Y., 2018, The remediation and safety utilization techniques for Cd contaminated farmland soil: a review, Shengtai Huanjing Xuebao (Ecology and Environmental Sciences), 27(7): 1377-1386. (孙丽娟, 秦秦, 宋科, 乔红霞, 薛永, 2018, 镉污染农田土壤修复技术及安全利用方法研究进展, 生态环境学报, 27(7): 1377-1386.)
[35] Tamás L., Mistrík I., Alemayehu A., Zelinová V., Bo?ová B., and Huttová J., 2015, Salicylic acid alleviates cadmium-induced stress responses through the inhibition of Cd-induced auxin-mediated reactive oxygen species production in barley root tips, J Plant Physiol, 173: 1-8.
[36] Wang Q.P., Chang M., and Lu L., 2020, Research progress of plant stress alleviation under heavy metal stress by plant growth promoting rhizobacteria, Heilongjiang Nongye Kexue (Heilongjiang Agricultural Sciences), 5:118-122.(王秋平, 常萌, 卢磊, 2020, 根际促生菌重金属胁迫下缓解植物应激的研究进展, 黑龙江农业科学, 5: 118-122.)
[37] Wang Y.C., Li J., Wang Y.F., Zhao X.L., Qiu C.S., Long S.H., and Guo Y., 2021, Study on different rhizosphere growth-promoting bacteria on agronomic characters and heavy metal cadmium and enrichment of flax, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 2021, 43(4): 194-204. (王元昌, 李洁, 王玉富, 赵信林, 邱财生, 龙松华, 郭媛, 2021, 不同根际促生菌对亚麻镉砷富集及植株生长的影响, 中国麻业科学, 2021, 43(4): 198-204.)
[38] Wang Y.F., Guo Y., Tang Q.M., Qiu C.S., Long S.H., Deng X., and Hao D.M., 2015, Study progress in flax remediation of soil contaminated by heavy metals and its application prospect, Zuowu Yanjiu (Crop Research), 29(4): 443-448. (王玉富, 郭媛, 汤清明, 邱财生, 龙松华, 邓欣, 郝冬梅, 2015, 亚麻修复重金属污染土壤的研究与应用. 作物研究, 29(4): 443-448.)
[39] Wu J.Y., and Mi N., 2022, Effect of heavy metals on antioxidant enzymes in plants, Zhejiang Nongye Kexue (Journal of Zhejiang Agricultural Sciences), 63(6): 1177-1181, 1304. (吴嘉煜, 米楠, 2022, 重金属对植物抗氧化酶影响研究进展, 浙江农业科学, 63(6): 1177-1181, 1304.)
[40] Wu Y.N., Pan G., Jiang H., Chang L., Huang S.Q., Tang H.J., and Luan M.B., Chen A.G., 2022, Advances in genome--wide association analysis of flax, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 44(2): 109-118. (伍叶娜, 潘根, 姜慧, 常丽, 黄思齐, 唐慧娟, 栾明宝, 陈安国, 2022, 亚麻全基因组关联分析研究进展, 中国麻业科学, 44(2): 109-118.)
[41] Xiang W., 2020, The effects of copper stress on flax germination, yield and physiological characteristics, Thesis for M.S., College of Plant Science, Huazhong Agricultural University, Supervisor:Peng D.X., pp.9-20. (向魏, 2020, 铜胁迫对亚麻萌发、产量及生理特性影响. 硕士学位论文, 华中农业大学植物科学技术学院, 导师: 彭定祥, pp.9-20)
[42] Xiang W., Peng D.X., and Liu L.J., 2022, The effects of copper stress on flax germination, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 44(4): 232-239.(向魏, 彭定祥, 刘立军,2022, 铜胁迫对亚麻萌发的影响, 中国麻业科学, 44(4): 232-239.)
[43] Xiong M.X., Wu D., Xu X.N., Zheng M.Y., and Xing T., 2021, Advances in toxic effects of soil heavy metal cadmium on higher plants, Shengtai Dulixue Bao (Asian Journal of Ecotoxicology), 2021, 16(6): 133-149.(熊敏先, 吴迪, 许向宁, 郑明阳, 邢涛, 2021, 土壤重金属镉对高等植物的毒性效应研究进展, 生态毒理学报, 16(6): 133-149.)
[44] Xu Y., Leng J., Jie Y.C., Sun Z.M., and Chen J.F., 2007, Studies on ecological adaptability of flax in Cd-polluted farmland, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 29(3): 136-138, 144. (许英, 冷鹃, 揭雨成, 孙志民, 陈建芳, 2007, 亚麻对镉污染农田适应性的初步研究, 中国麻业科学, 29(3): 136-138, 144.)
[45] Xu Y., Wei X.H., and Li B.B., 2012, Effects of exogenous nitric oxide on antioxidant capability of seeding leaves in linseed under Cd stress, Gansu Nongye Daxue Xuebao (Journal of Gansu Agricultural University), 47(5): 45-49, 55. (徐严, 魏小红, 李兵兵, 2012, 外源NO对重金属Cd胁迫下亚麻幼苗叶片抗氧化能力的影响, 甘肃农业大学学报, 47(5): 45-49, 55.)
[46] Yang Y.D., 2021, The comparison of remediation potential of multi season economic crop rotation systems to cadmium-polluted in farmland, Thesis for M.S., College of Resources and Environment, Hunan Agricultural University, Supervisor: Zeng Q.R., pp.9-12. (杨耀东, 2021, 多季经济作物轮作对镉污染农田修复潜力的比较, 硕士学位论文, 湖南农业大学资源环境学院, 导师: 曾清如, pp.9-12.)
[47] Zhang X.Y., Ye Z.B., and Zhang Y.Y., 2021, Advances in physiological and molecular mechanism of plant response to cadmium stress, Zhiwu Shengli Xuebao (Plant Physiology Journal), 57(7): 1437-1450. (张星雨, 叶志彪, 张余洋, 2021, 植物响应镉胁迫的生理与分子机制研究进展, 植物生理学报, 57(7): 1437-1450.)
[48] Zhao D.X., Dong Y.R., Sun J.S., Geng B., Wang Z.H., Guo H.E., and Wang X.Y., 2020, Effects of NaCl stress on seed germination, seedling growth and Na+/H+ antiporter protein gene expression of Linum usitatissimum L, Shandong Nongye Kexue (Shandong Agricultural Sciences), 52(7): 40-45.(赵东晓, 董亚茹, 孙景诗, 耿兵, 王照红, 郭洪恩, 王向誉, 2020, NaCl胁迫对胡麻种子萌发、幼苗生长及Na~+/H~+逆向转运蛋白基因表达的影响, 山东农业科学, 52(7): 40-45.)
[49] Zhao X.L., Wang H., Qiu H.J., Qiu C.S., Long S.H., Wang Y.F.,and Guo Y., 2022, Effects of cadmium treatment on the flax growth and distribution of cadmium in flax, Hunan Nongye Daxue (Journal of Hunan Agricultural University (Natural Sciences Edition)), 2022, 48(1): 82–86.(赵信林, 王慧, 邱化蛟, 邱财生, 龙松华, 王玉富, 郭媛, 2022, 镉处理对亚麻生长发育的影响及镉在亚麻体内的分布, 湖南农业大学学报(自然科学版), 2022, 48(1): 82-86.)
[50] Zhao X.L., Wei X.Y., Guo Y., Qiu H.J., Qiu C.S., Long S.H., and Wang Y.F., 2020, Effects of foliar application of organic acids on the growth and cadmium-remediation efficiency of flax, Zhongguo Maye Kexue (Plant Fiber Sciences in China), 42(6): 257-262. (赵信林, 韦秀叶, 郭媛, 邱化蛟, 邱财生, 龙松华, 王玉富, 2020, 叶面喷施有机酸对亚麻生长发育及镉修复能力的影响, 中国麻业科学, 42(6): 257-262.)
文章摘自:刘琴,安霞,杜光辉.亚麻在修复重金属污染土壤上的运用[J/OL].分子植物育种:1-9[2023-10-10].http://kns.cnki.net/kcms/detail/46.1068.S.20230918.1628.018.html
