摘 要:罗布麻类是具有强抗逆性、高耐受性的典型植物,更是具有开发利用价值的经济作物.本研究应用ENMTools工具优化的最大熵模型(MaxEnt)模拟罗布麻(Apocynum)类的白麻(Apocynum pictum (Sohnenk.) Baill)、大叶白麻(Apocynum hendersonii (Hook.f.) Woodson.)和罗布红麻(Apocynum venetum L.)3个物种.分析了在当前气候条件下及RCP4.5排量模式下,未来2个时段(2041−2050年和2061−2070年)的潜在空间分布格局和影响各物种分布的主要环境因子对物种的影响.结果显示:1)3个物种的地理分布受到不同的主导环境因素影响.最湿季度降水量、降水量季节性变化、海拔是影响白麻分布的主导环境因子;影响大叶白麻分布的主要环境因子包括昼夜温差月均值、最热月份最高温度、最湿季度降水量、坡向和表层(0~30cm)砂粒含量;对于罗布红麻来讲,海拔、年温变化范围、最湿季度降水量、降水量季节性变化是影响其分布的关键因素.2)3个物种适生区在地理位置分布上存在明显差异.3)白麻和大叶白麻之间存在生态位重叠度,而罗布红麻与其他2个物种的生态位重叠较低.未来,白麻和大叶白麻的分布中心向高纬度、高海拔地区移动,而罗布红麻的分布中心呈向北移动趋势.这些研究结果为罗布麻类植物引种栽培和保护提供理论指导,并为探究气候变化对物种分布的影响提供参考.
关键词:罗布麻;MaxEnt;潜在分布格局;生态适宜性;ENMTools
罗布麻类植物隶属夹竹桃科(Apocynaceae)罗布麻属(Apocynum),是珍稀的野生植物资源.我国分类学家将罗布麻划分为2个属3个种,即白麻(A.pictum)(又称紫斑中花罗布麻)、大叶白麻(A.hendersonii)(又称大花罗布麻)和罗布红麻(A.venetum L.)(又称罗布麻)[1].罗布麻在药用方面具有悠久历史,其根、茎、叶、花全株均可入药,被认为具有强心抗炎、止咳平喘、降低血脂等功效[2].此外,罗布麻类植物还具有抗盐碱、耐干旱、抗风沙、耐病虫害的植物学特征,因此被认为是一种具有较高生态价值的荒漠先锋植物,能够改善气候条件、涵养水土、防风固沙、改良土壤,促进盐碱地内部形成良性循环.相比于其他植物,罗布麻在荒漠化地区的生存能力更强,具有不可替代的优势.但是,由于气候变化、生态环境恶化、人工过度采挖等因素的影响,罗布麻资源面积急剧缩减[2−3].目前,针对罗布麻的研究多集中在小范围的理化成分分析.杨会枫等[4]利用最大熵(MaxEnt)模型默认参数,预测了罗布麻在未来50年的潜在分布和主导环境因子,而并未将罗布麻类3个物种区分开来.因此,分区研究罗布麻类3种植物在中国的潜在适宜性分布,探讨其与环境因子间适应机制,预测未来潜在适生区变化,能够为现有罗布麻资源的有效保护和人工引种提供有效理论依据,对研究干旱地区气候变化对植物迁移的影响,维持生态可持续发展有重要意义.
政府间气候变化专门委员会(IPCC)第6次评估报告(AR6)显示,自1850−1900年以来,全球地表平均温度已上升约1℃,并指出从未来20年的平均温度变化来看,全球升温预计将达到或超过1.5℃[5].全球升温势必影响物种分布.有研究表明,平均每10年,陆地类群向极地移动17km[6],海洋类群平均每10年向极地移动72km[7],山坡上的陆生物种为躲避高温而向上移动[6].更有学者指出,随着未来全球气温升高、物种灭绝的风险将会增大,目前全球约1/6的物种因气候变化面临灭绝风险[8].因此,为降低罗布麻灭绝风险,合理进行人工引种栽培,有必要研究罗布麻地理分布格局对未来气候变化的响应.
MaxEnt模型是具有较高准确性的物种分布模型之一,其利用物种分布数据和环境数据,根据特定算法模拟物种的分布,并反映物种对栖息地的偏好程度[9].MaxEnt模型通过使用刀切法(Jackknife)来判断环境因素的重要性,并定量描述环境因素对物种栖息地的影响.模型的准确性通过受试者工作特征曲线(ROC)的曲线下面积(AUC)进行验证.研究表明,相对于其他物种分布模型,MaxEnt模型对小样本数据的预测效果更好、准确性更高[10].然而,不同物种的生理特征、生态适应能力各不相同,仅使用MaxEnt默认参数获得预测结果,其准确性已不能满足实际需求,需要探寻适应于各物种的最佳参数以优化模型.
本研究通过野外实地调研结合大量的考察记录资料,采用基于R语言构建的生态地理学ENMTools(V3.4.4)工具,对MaxEnt模型参数进行优化,模拟罗布麻类白麻、大叶白麻和罗布红麻3种植物在中国的地理分布.以期明确影响罗布麻3个物种分布范围的重要环境因素;利用模型模拟当前和未来气候条件下3个物种的分布变化趋势,探究气候变化对物种分布的影响;利用未来气候条件量化适宜生境范围变化的空间模式,为罗布麻的实际引种栽培提供科学依据.
1 数据与方法
1.1 物种分布数据搜集与筛选
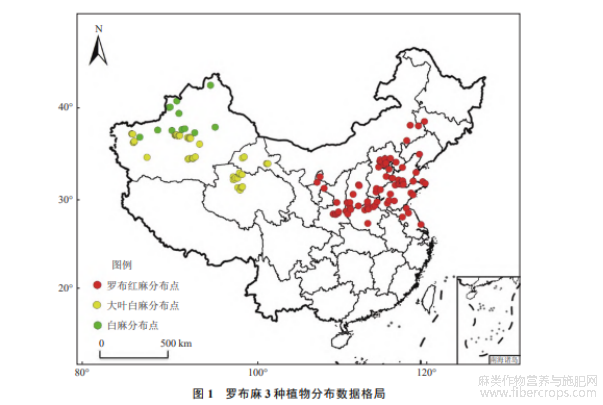
MaxEnt模型模拟所需数据包括物种分布数据和环境变量数据,其中物种分布数据指的是分布点的经纬度信息.罗布麻类3种植物数据通过下列方式获得:1)野外实地调研,于2021年夏季进行全国调研,调研位点遍及全国罗布麻类历史分布区,用GPS记录经纬度信息;2)数据库查询,本研究所查询数据库包括“中国植物标本馆(CVH,https://www.cvh.ac.cn/)”和“全球物种多样性信息库(GBIF,https://www.gbif.org/)”;3)文献查询,检索出近50年来罗布麻相关的期刊论文.本研究共搜集到白麻样点20个、大叶白麻样点70个、罗布红麻样点85个.
使用ENMTools软件筛选分布点,以便在每5′栅格只保留1个分布点.最后,用于导入模型的分布点数量分别为白麻12个、大叶白麻48个、罗布红麻78个(图1).
1.2 环境变量数据
1.2.1 环境变量数据来源
本研究所需的环境变量包括气象因子、地形因子和土壤因子3类.气候数据均来自于WorldClim环境数据库(http://www.worldclim.org/),空间分辨率5′,包括当前(1970−2000年)和未来(2041−2050年,2061−2070年)的气候数据.其中,未来数据采用CCSM4模式中的RCP4.5情景,该情景下温室气体排放量与当前状况(1970−2000年)大致相同.本研究假设地形因子和土壤因子在21世纪保持不变.地形因子包括海拔、坡向和坡度,在ArcGIS10.8中利用DEM高程数据(https://www.gscloud.cn/)提取上述地形参数.土壤因子从中国科学院资源环境科学数据中心(https://www.resdc.cn/)下载,罗布麻多生长于砂质土壤地区[11],故选择的土壤因子包括表层和底层砂粒、黏粒含量.
1.2.2 环境变量筛选
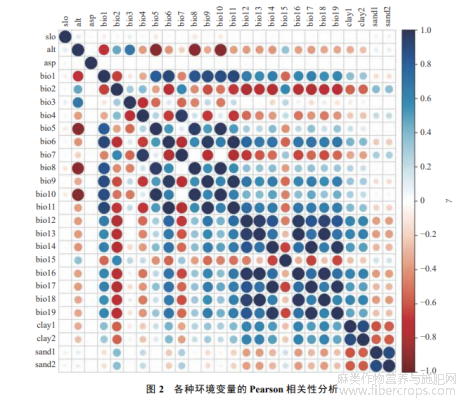
环境变量是构建生态位模型的重要参数,使用过多的环境变量会造成过拟合,降低模型的准确性[12−13].因此,需要排除相关性高的环境变量.本研究使用ENMTools.pl软件对备选变量进行相关性分析,当Pearson相关系数|r|>0.8时,定义为高度相关变量[14](图2).
1.3 模型参数优化
使用Kuenm R包(https://github.com/marlonecobos/kuenm)来优化MaxEnt模型参数[9].特征组合(FC)参数和倍频设置(RM)是对使用MaxEnt构建物种分布模型重要的2个参数.在建模中,将RM值设置为0.1~4.0,每次增加0.1,采用29个特征组合,共评估了1160个候选模型.信息量准则(AIC)是衡量模型拟合优良性的标准之一,模型选择基于统计显著性(部分ROC)、预测能力(低遗漏率)和修正的AIC(AICc)值,按优先顺序排列:首先,筛选以保留具有统计学意义的模型;其次,使用遗漏率标准减少模型集;最后,选择delta AICc值最小的模型,认为是最佳模型.
1.4 物种分布模型的构建与评估
本研究利用MaxEnt得到的每个物种的模型,具有不同的正则化乘数和特征类参数.运行模型时,75%的样本用于训练,25%用于模型验证.模型重复运行10次,设置最大迭代次数5000次、最大背景点数10000个.通过刀切法评估每个环境变量的相对贡献[16−17].ROC的AUC方法用于评估MaxEnt模型的准确度.理论上,AUC值为0.5~0.7时,效果较差;0.7~0.9时,效果一般;>0.9效果较好[18].
1.5 生境适宜性划分
MaxEnt输出的每个栅格中的概率值在0~1之间,可以表述为相对适宜性.本研究所使用的中国地图矢量数据均来自于标准地图服务系统(http://bzdt.ch.mnr.gov.cn).利用ArcGIS10.8对模型结果进行可视化表达,依据自然断点法结合物种分布规律将罗布麻类植物潜在适生区划分为非适生区(适生值(0,0.1])、低适生区((0.1,0.3])、中适生区((0.3,0.6])、高适生区((0.6,1.0])[19]4个等级.
1.6 物种适生区空间格局变化
1.6.1 分布面积及分布中心变化
1)不同物种各时期面积.将重分类后的栅格文件导入ArcGIS10.8,使用“AttributeTable”功能,获得各适生等级的栅格数量“COUNT”,分别计算各适生区占比和适生区面积.
2)分布中心变化的获得.采用SDMToolbox(http://www.sdmtoolbox.org/downloads)工具包,将3个物种不同时期的预测结果文件转换为二进制的分布文件,然后选择“SDMTools”模块中“UniversalTools”子目录,“CentroidChanges(Lines)”用于计算不同时期预测分布的几何中心位移情况,检测分布区的总体变迁趋势[20].
1.6.2 生态位重叠度和地理分布重叠度
将MaxEnt模型的预测结果导入ENMTools(V1.4.4)计算生态位重叠度和地理分布重叠度.本研究采用ENMTools中的D测量法计算生态位重叠度
![]()
式中:Oij表示物种i和物种j之间的生态位重叠度,Pia和Pja分别代表物种i和物种j利用资源a的个体数.理论上,生态位重叠度≥0.5视为重叠度高,0.3~
2 结果与分析
2.1 模型优化及准确性评估
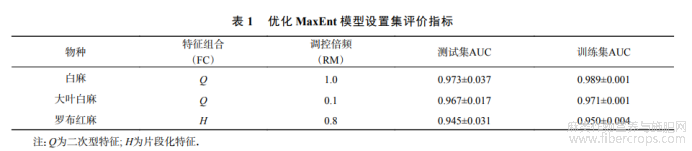
基于罗布麻类植物分布点和10个生态因子,分别采用优化后的MaxEnt模型进行潜在分布预测.经过模型参数优化,delta.AICc达到最小(0),故选择白麻最佳参数模式的FC=Q(二次型特征),RM=1.大叶白麻最佳参数模式的FC为Q,RM为0.1;罗布红麻最佳参数模式的FC为H(片段化特征),RM为0.8(表1).本研究选取以上3种参数结果作为建模的参数设置.在优化参数下,MaxEnt模型预测结果显示3个物种的平均训练集AUC均>0.9(表1),这表明MaxEnt模型预测到的中国罗布麻类植物潜在分布区具有较好的精度,拟合程度高,预测结果并非随机.
2.2 环境因子影响分析
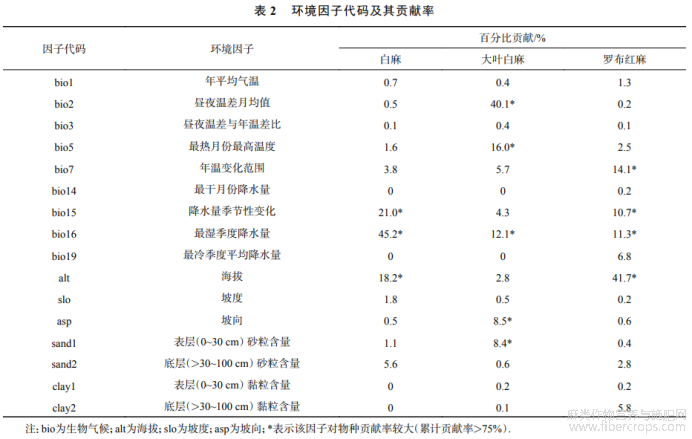
将通过环境因子筛选的16个环境因子输入MaxEnt模型(表2),利用刀切法确定主导环境因子,根据大小顺序,认定累计贡献率达到75%以上的因子为主导环境因子[22].结果表明:白麻分布模型的主导环境因子为最湿季度降水量、降水量季节性变化、海拔,累计贡献率达84.4%(表2).大叶白麻的主导环境因子为昼夜温差月均值、最热月份最高温度、最湿季度降水量、坡向和表层(0~30cm)砂粒含量,累计贡献率达85.1%.罗布红麻的主导环境因子是海拔、年温变化范围、最湿季度降水量、降水量季节性变化(表2),累计贡献率达77.8%.
基于上述结果,本研究将主导环境因子作为限制因子,分别分析适宜白麻、大叶白麻和罗布红麻生长及分布的环境特征.
2.3 适宜生境条件
基于各生态因子响应曲线(图3)可知,当适生程度>0.5时,其所对应的环境变量数值较为适宜罗布麻3个品种生长及分布.
2.3.1 白麻适宜生境条件
刀切法结果表明,影响白麻地理分布的生态因子为最湿季度降水量、降水量季节性变化、海拔.海拔>925m时,白麻的适生程度65.45mm时,白麻适生程度78.79mm时,白麻适生程度<0.5,随着最湿季度降水量增加,白麻适生程度急剧下降,故白麻适宜生长的最湿季度降水量为3.88~78.79mm(图3).因此,白麻适宜分布在海拔较低、降水量季节性变化不大且干旱的地区.
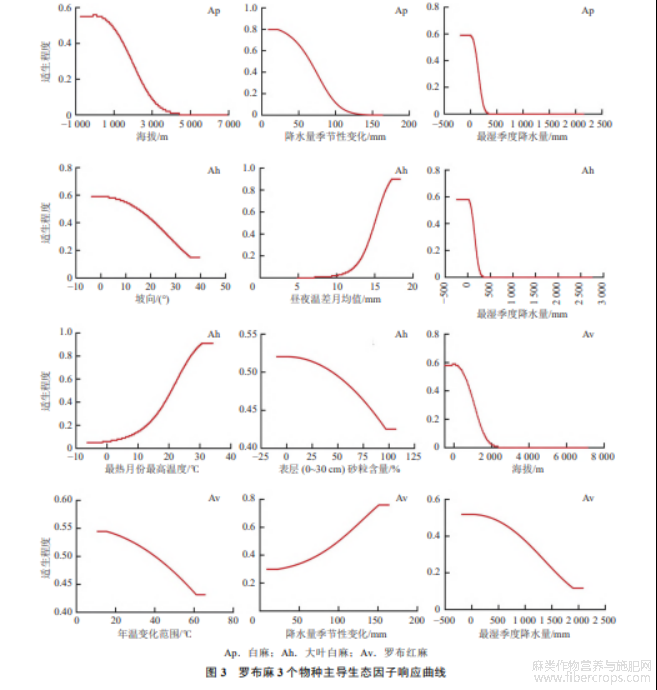
2.3.2 大叶白麻适宜生境条件
影响大叶白麻地理分布的主导环境因子是昼夜温差月均值、最热月份最高温度、最湿季度降水量、坡向、表层砂粒含量.当坡向为15°时,大叶白麻的适生程度为0.5,随着坡向的增加,适生程度缓慢下降,因此适宜大叶白麻生长的坡向为0~15°.当昼夜温差月均值<10℃时,大叶白麻适生程度趋近于0,>14.5℃后适生程度>0.5,故昼夜温差月均值的适宜性范围为14.5~18.3℃.当最湿季度降水量为3.01~86.81mm、最热月份最高温度为20.71~34.18℃、表层(0~30cm)砂粒含量为2.1%~44.8%时,大叶白麻适生程度较高.综上所述,大叶白麻适宜分布于地势平缓、昼夜温差较大、较为干旱、夏季炎热且表层土壤砂粒含量较低的地区.
2.3.3 罗布红麻适宜生境条件
刀切法结果显示,影响罗布红麻分布的主导环境因子包括年温变化范围、降水量季节性变化、最湿季度降水量和海拔.在海拔0.5,适宜生长,但随着海拔升高,适生程度下降,因此,罗布红麻海拔适宜分布范围为−154~600m.年温变化范围为10.72~40.07℃时,罗布红麻的适生概率最高,适生概率随年温变化范围升高而降低.降水量季节性变化为100.12~163.91mm时,罗布红麻适生概率最高.罗布红麻在最湿季度降水量1.97~356.02mm范围内适宜生长.因此,罗布红麻适宜分布在海拔较低、年温变化范围适中、降水量季节性变化较大且轻度干旱地区.
2.4 潜在适生区及未来变化
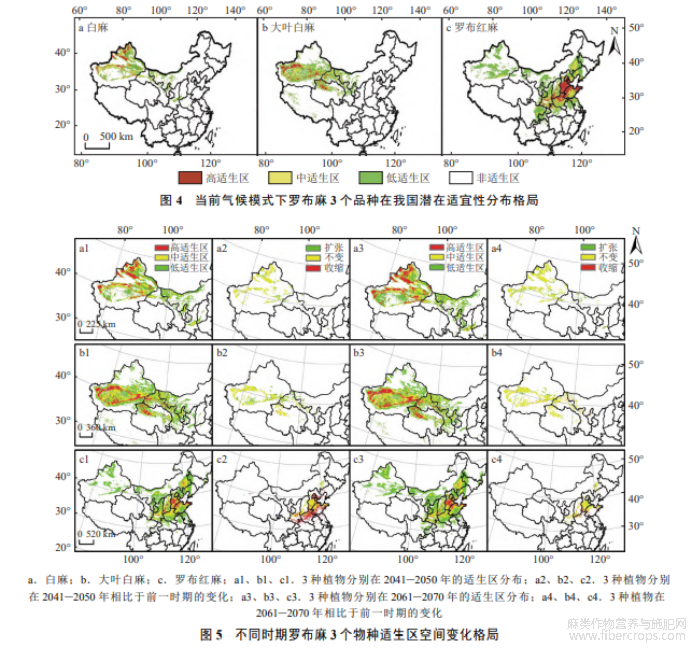
2.4.1 当前气候条件下适生区分布
当前,白麻适生区主要在我国西北部,面积为6.14×105km2,约占国土面积的6.40%.其中,高适生区分布于新疆乌鲁木齐、阿勒泰等地(图4-a),面积4.93×104km2,约占国土面积的0.51%;中适生区面积1.77×105km2,约占国土面积的1.84%;低适生区面积为3.87×105km2,约占国土面积的4.03%.
大叶白麻潜在分布适宜区包括新疆中部、青海北部、内蒙古西部等地区(图4-b),总面积为1.38×106km2,占国土面积的14.37%.高适生区分布较为分散(图4-b),面积为1.60×105km2,约占国土面积的1.67%;中适生区约占国土面积的4.29%,面积为4.12×105km2,主要围绕高适生区分布;低适生区沿高、中适生区发散分布,分布面积8.08×105km2,约占国土总面积的8.41%.
罗布红麻适生区分布最广,面积高达2.08×106km2,约占国土面积的21.67%;高适生区主要分布在河北、山东等地,面积约为2.79×105km2,占国土面积的2.91%;中适生区面积约为5.37×105km2,占国土面积的5.59%;低适生区环绕中、高适生区,分布面积约为1.26×106km2,约占13.13%(图4-c).
2.4.2 适宜分布区未来变化趋势
MaxEnt模型预测显示,白麻适宜分布区未来呈扩张趋势(图5).在2041−2050年气候条件下,扩张范围主要发生在新疆克拉玛依、陕西西安等地,新增面积约4.54×104km2,约占当前模式适宜生境的7.39%;收缩范围主要位于山西运城等地,收缩面积约202.62km2,约占当前适宜生境面积的0.03%.在2061−2070年气候模式下,与前一时期相比,白麻潜在分布范围保持扩张趋势,扩张面积约7.19×104km2,占2041−2050年时期适生区面积的11.71%
大叶白麻的未来潜在适宜分布区总体呈扩张趋势,但在部分地区存在局部收缩(图5).在2041−2050年时期,潜在适宜分布区面积扩张主要发生在内蒙古阿拉善右旗、青海西宁等地,扩张面积约1.04×105km2,相比当前气候模式增长7.53%,面积收缩区域主要位于青海格尔木等地,收缩面积约2938km2.在2061−2070年时期,相比于2041−2050年时期,大叶白麻的潜在适宜分布范围保持扩张趋势,新增面积约3.93×104km2,占2041−2050年时期适生区面积的2.85%,收缩面积约9219km2,占比0.67%.高适生区面积占比有所增加,中、低适生区面积占比有所降低.新增和收缩区域在2个时期基本一致.
未来罗布红麻分布区局部收缩严重.在2050年时期,潜在适生区收缩面积约2.06×105km2,占当前适生区面积的9.90%,各适生区占比均减少.收缩区域主要位于河南、山东等地.2061−2070年时期,潜在适宜分布区收缩略有好转,扩张面积约2.85×104km2,占2041−2050年适生区面积的1.37%,扩张区域主要位于北京、天津等省区,收缩区域面积约3.67×104km2,占比1.37%,各适生区面积变化仅高适生区占比略有降低,中适生区占比由5.59%增加至5.81%,低适生区占比由13.13%增加至14.67%(图5).
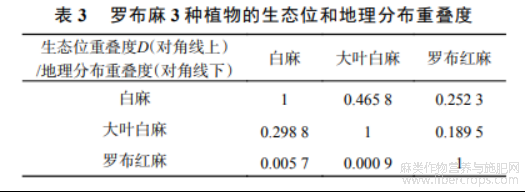
2.5 生态位重叠度和分布中心转移
表3显示了罗布麻类3个物种之间的生态位重叠度和地理分布重叠度,3个物种间白麻与大叶白麻表现出适中的生态位重叠度(D=0.4658),罗布红麻和白麻、大叶白麻之间生态位重叠度均<0.3.此外,除白麻和大叶白麻,其余物种间地理分布重叠度均较低.
白麻在当前气候条件下的分布中心位于新疆维吾尔自治区吐鲁番市托克逊县库加依镇南部,海拔690m,坐标北纬43°08′、东经85°17′.2041−2050年的分布中心位于新疆巴音郭楞蒙古自治州和静县托拉特郭勒北部,海拔2585m,移动约68.6km.2061−2070年分布中心位于新疆巴州和静县北部山区巴伦台镇,海拔3558m,移动62.5km(图6-a).总体来看,白麻分布中心向高海拔地区和东南方向移动.
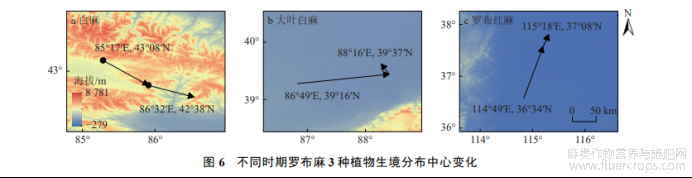
大叶白麻的当前分布中心位于新疆维吾尔自治区巴音郭楞蒙古自治州若羌县瓦石峡镇,海拔约415m,坐标为北纬39°16′、东经86°49′.在2041−2050年气候条件下,分布中心移动至若羌县喀拉布然库勒西北部,海拔805m,距当前气候条件的分布中心约154.2km.2061−2070年气候模式下分布中心海拔810m,坐标为北纬39°37′、东经88°16′,相比于2041−2050年气候条件下,分布中心移动22.3km(图6-b).大叶白麻分布中心的垂直梯度呈向高海拔地区移动趋势,水平梯度则向东北方向移动.
罗布红麻分布中心在水平梯度向东北方向迁移,垂直梯度呈先降后升趋势.在当前气候条件下,分布中心位于河北省邯郸市肥乡区肥乡镇南西落堡村,海拔50m,坐标北纬36°34′、东经114°49′.在2041−2050年时期,分布中心位于河北省邢台市沙河市柴关乡高庄村,海拔29m,距当前分布中心约108.6km.2061−2070年时期,分布中心位于河北省石家庄市辛集市马庄乡西谢村北部,海拔33m,分布中心移动25.7km(图6-c).
3 讨论
白麻的地理分布受最湿季度降水量、降水量季节性变化、海拔等因素影响(表2),这与汤博[23]的结论相符.最湿季度降水量是影响白麻分布的最重要因素,表明降水量对白麻生长至关重要,最湿季度降雨量3.88~78.79mm,该降水条件表明白麻适宜生长在较为干旱的环境中.孙丽君[24]的研究表明,白麻具有更强的耐旱性和耐盐性能力,生境相比红麻更为干旱,与本文结果一致.白麻潜在分布区主要集中于我国西北内陆地区,这些区域大部分土壤盐碱化程度很高[24].因此,在西北内陆荒漠、半荒漠及盐渍地区开展白麻集约化种植,可以缓解土地荒漠化趋势,构建良好的生态环境.
大叶白麻的适生程度主要受昼夜温差月均值、最热月份最高温度,最湿季度降水量、坡向和表层(0~30cm)砂粒含量因子的影响(图3).大叶白麻的这种生态区位特征与其生理特性密切相关:适度干旱会促使大叶白麻的根系延长,分蘖增多,利用其发达的根系广泛吸收水分,使其适应干旱的环境.此外,大叶白麻具有盐生植物和旱生植物所特有的高渗透压细胞,可在盐分含量大的土壤中吸收水分[25−26].较大的昼夜温差有利于光合作用,而夜间较低的温度可以减少能量消耗[27].有研究表明,大叶白麻在一定程度上受地下水位供给限制[4].大叶白麻在新疆、青海和甘肃等省区内主要存在于盐碱荒地和沙漠边缘[1],这可能与地下水位的分布具有密切关系.本研究将青海柴达木盆地也划分为大叶白麻适生区,在该地区种植大叶白麻有助于改善当地农业产业结构,促进经济发展.
罗布红麻适宜生长在海拔较低、年温变化范围适中、降水量季节性变化较大且轻度干旱地区.前人的研究表明,温度、水分、海拔是影响罗布红麻分布的主要非生物因素[26].罗布红麻适应能力强,能在极端高温、低温和干旱条件下生存.其具有独特的适应干旱机制,包括保护酶体系和渗透调节物质[28],使其能够在中度干旱条件下生长[29−30],并在10~25℃范围内与温度呈正相关[31].罗布红麻主产于东北、西北、华北和黄河流域等省区[32],但其实际分布可能延伸到基础生态位以外的区域[9,33−35].模型预测其适生区面积高达2.08×106km2,约占我国国土面积的21.67%.然而,由于人类活动的影响,实际适生区可能小于模拟结果.
随着全球气候变暖,陆生物种往往通过移动位置来适应变化.为了追求适宜的环境,物种倾向于迁徙到较冷的高海拔或高纬度地区[7],而另一部分物种由于其自身的生理特征导致分布响应滞后于气候变化[36].随着气候变暖、物种间相互作用,物种分布范围会趋向于冷边缘范围(高纬度和高海拔)拓展和暖边缘范围(低纬度和低海拔)收缩[37].尽管物种可能通过表型可塑性或自然选择来适应不断变化的气候[38],但物种的适应能力都是有限的[39],对于已经达到耐受极限的物种来说,其适应能力不可能再增加[40].研究物种分布边缘收缩具有重要意义,当原有分布区的最低纬度或最低海拔区域不再存在该物种,这表明物种未能充分改变其生态位以适应新的环境条件,表明该物种可能正在走向局部灭绝[41].本研究采用RCP4.5排放情景下的未来气候数据进行预测,结果显示,白麻和大叶白麻的分布中心向高纬度和高海拔地区移动,而罗布红麻的分布中心向北部移动.罗布红麻这种分布中心移动滞后于气候变化的情况,或许与其地理分布位置有关,罗布红麻高适生区多分布于沿海地区,全球气候变暖不仅使海平面加速上升,还有观测表明中国东部沿海地区物候期提前,干旱范围扩大,这一系列影响都有可能导致罗布红麻在未来向北部迁移.此外,白麻和大叶白麻适生区面积收缩,这表明即使较小的气候变化也足以导致植物的局部灭绝,且强抗逆性的罗布红麻类植物也无法避免这种趋势.全球变暖情况未能改善,并且随着碳排放的增加进一步恶化,更多的物种可能会因难以快速响应气候变化而面临灭绝风险.
本研究仅考虑了气候变化和环境因子对物种分布的影响,未考虑物种间相互作用和人类活动的影响.因为特定的模型分析中不可能考虑到所有因素的影响,物种分布模型的模拟结果仅为理想状态的潜在分布范围[42].此外,由于相关影响因素数据,如土地利用变化和人类活动等难以获得,因此如何将这些因素纳入模型,进一步提高模型模拟结果的准确度,也是未来物种分布模型研究需要探讨的方向.
4 结论
本研究利用ENMTools优化MaxEnt模型参数,预测了罗布麻类3个物种在当前及未来气候模式下的潜在适生区,并分析了影响它们分布的主要环境因子和适生范围,主要结论如下.
1)最湿季度降水量、降水量季节性变化、海拔是影响白麻分布的主导环境因子;昼夜温差月均值、最热月份最高温度、最湿季度降水量、坡向和表层(0~30cm)砂粒含量是影响大叶白麻分布的主导环境因子;海拔、年温变化范围、最湿季度降水量、降水量季节性变化是影响罗布红麻分布的主导环境因子.
2)未来气候条件下,白麻、大叶白麻分布中心有向高纬度、高海拔地区转移趋势;罗布红麻分布中心则向北部转移,海拔变化并不明显.
3)白麻、大叶白麻和罗布红麻的潜在适生区将在未来继续扩张,分别集中在我国西北部的新疆中部、甘肃、青海北部和内蒙古西部边缘,以及河北、山东沿海及沿黄河分布地区、河南北部地区.
上述结论对栽培引种提供理论指导,同时也说明较小的气候变化也会威胁强耐受性植物的生存.因此减少碳排量、缓解全球变暖趋势对维护生物多样性具有重要意义.
参考文献
[1] 中国科学院中国植物志编辑委员会. 中国植物志:第63卷 [M]. 北京:科学出版社,1977
[2] 谭燕玲,徐原,朱静. 罗布麻的性能与现状研究[J]. 轻纺工业与技术,2021,50(1):16
[3] 苏前,邱连勇. 新疆罗布麻资源概况[J]. 农业灾害研究, 2015,5(6):61
[4] 杨会枫,郑江华,贾晓光,等. 气候变化下罗布麻潜在地理分布区预测[J]. 中国中药杂志,2017,42(6):1119
[5] ALLAN R P, HAWKINS E, BELLELLOUIN N, et al. IPCC, 2021: summary for policymakers[M]. Cambridge County,UK:Cambridge University Press,2021
[6] CHEN I C,HILL J K,OHLEMüLLER R,et al. Rapid range shifts of species associated with high levels of climate warming[J]. Science,2011,333(6045):1024
[7] POLOCZANSKA E S,BROWN C J, SYDEMAN W J, et al. Global imprint of climate change on marine life[J]. Nature Climate Change,2013,3(10):919
[8] ZHANG K L,YAO L J,MENG J S,et al. MaxEnt modeling for predicting the potential geographical distribution of two peony species under climate change[J]. Science of the Total Environment,2018,634:1326
[9] COBOS M E,PETERSON A T,BARVE N,et al. Kuenm: an R package for detailed development of ecological niche models using MaxEnt[J]. PeerJ,2019,7:e6281
[10] PEARSON R G, RAXWORTHY C J, NAKAMURA M, et al. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar[J]. Journal of Biogeography,2006,34(1):102
[11] ZHU G P,PETERSEN M J,BU W J. Selecting biological meaningful environmental dimensions of low discrepancy among ranges to predict potential distribution of bean plataspid invasion[J]. PLoS One,2012,7(9):e46247
[12] 雷菁清. 黄河上游谷地罗布麻耕地土壤质量水平及产品品质特征研究[D]. 西宁:青海大学,2021
[13] SAUPE E E,BARVE V,MYERS C E,et al. Variation in niche and distribution model performance: the need for a priori assessment of key causal factors[J]. Ecological Modelling,2012,237/238:11
[14] KONG F,TANG L,HE H,et al. Assessing the impact of climate change on the distribution of Osmanthus fragrans using MaxEnt[J]. Environmental Science and Pollution Research,2021,28(26):34655
[15] ZHAO G H, CUI X Y, SUN J J, et al. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models[J]. Ecological Indicators,2021,132:108256
[16] PHILLIPS S J, ANDERSON R P, DUDíK M, et al. Opening the black box: an open-source release of MaxEnt[J]. Ecography,2017,40(7):887
[17] PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions[J]. Ecological Modelling, 2006, 190(3/4): 231
[18] YANG J T, HUANG Y, JIANG X, et al. Potential geographical distribution of the edangred plant Isoetes under human activities using MaxEnt and GARP[J]. Global Ecology and Conservation,2022,38:e02186
[19] 刘超,霍宏亮,田路明,等. 不同气候情景下木梨潜在地 理 分 布 格 局 变 化 的 预 测 [J]. 应 用 生 态 学 报 , 2020, 31(12):4073
[20] BROWN J L, BENNETT J R, FRENCH C M. SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses[J]. PeerJ,2017,5:e4095
[21] WARREN D L,GLOR R E,TURELLI M. ENMTools:a toolbox for comparative studies of environmental niche models[J]. Ecography,2010,35:607
[22] 孟影,马姜明,王永琪,等. 基于MaxEnt模型的檵木分布 格局模拟[J]. 生态学报,2020,40(22):8287
[23] 汤博. 新疆地区罗布白麻(Poacynum hendersonii)的生态 适应性特征研究[D]. 北京:中国环境科学研究院,2011
[24] 孙丽君. 不同环境条件下罗布麻和白麻光合特性研究 [D]. 乌鲁木齐:新疆大学,2011
[25] 刘建新,王鑫,王凤琴. 水分胁迫对苜蓿幼苗渗透调节物 质积累和保护酶活性的影响[J]. 草业科学,2005,22(3): 18
[26] 李霞,马永清,宋玉霞,等. 肉苁蓉寄生对梭梭幼苗保护 酶活性及渗透调节物质的影响[J]. 生态学杂志,2009, 28(8):1531
[27] 石秋梅,邓翻云,吴敏言,等. 罗布麻和大叶白麻种子萌 发及幼苗生长耐盐性研究[J]. 北方园艺,2014(12):128
[28] 徐振朋,宛涛,蔡萍,等. PEG模拟干旱胁迫对罗布麻种 子萌发及生理特性的影响[J]. 中国草地学报, 2015, 37(5):75
[29] 宁建凤,郑青松,杨少海,等. 高盐胁迫对罗布麻生长及 离子平衡的影响[J]. 应用生态学报,2010,21(2):325
[30] 胡瑞林,林木兰,董正钧. 罗布麻生长发育规律的探讨:Ⅰ. 罗布麻种子发芽与温度的关系[J]. 中国野生植物资 源,1998,17(3):35
[31] 胡瑞林,钱学射,肖永绥,等. 罗布麻生长发育规律的探 讨:Ⅲ. 罗布麻茎的生长规律[J]. 中国野生植物资源, 1998,17(3):37
[32] 刘起棠,张卫明,肖正春,等. 我国中东部地区罗布红麻 资源濒危状况探讨[J]. 中国野生植物资源,2009,28(3): 9
[33] WALDEN-SCHREINER C,LEUNG Y F,KUHN T,et al. Environmental and managerial factors associated with pack stock distribution in high elevation meadows: case study from Yosemite National Park[J]. Journal of Environmental Management,2017,193:52
[34] WAN G Z, WANG L, JIN L, et al. Evaluation of environmental factors affecting the quality of Codonopsis pilosula based on chromatographic fingerprint and MaxEnt model[J]. Industrial Crops and Products, 2021, 170: 113783
[35] HEBBAR K B, ABHIN P S, SANJO JOSE V, et al. Predicting the potential suitable climate for coconut (Cocos nucifera L. ) cultivation in India under climate change scenarios using the MaxEnt model[J]. Plants (Basel, Switzerland),2022,11(6):731
[36] LENOIR J,SVENNING J C. Climate-related range shifts: a global multidimensional synthesis and new research directions[J]. Ecography,2015,38(1):15
[37] LENOIR J, GÉGOUT J C, GUISAN A, et al. Going against the flow: potential mechanisms for unexpected downslope range shifts in a warming climate[J]. Ecography,2010:295
[38] VALLADARES F,MATESANZ S,GUILHAUMON F,et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change[J]. Ecology Letters,2014,17(11):1351
[39] WILLIAMS S E,SHOO L P,ISAAC J L,et al. Towards an integrated framework for assessing the vulnerability of species to climate change[J]. PLoS Biology,2008,6(12): 2621
[40] ARAúJO M B, FERRI-YáñEZ F, BOZINOVIC F, et al. Heat freezes niche evolution[J]. Ecology Letters, 2013, 16(9):1206
[41] WIENS J J. Climate-related local extinctions are already widespread among plant and animal species[J]. PLoS Biology,2016,14(12):e2001104
[42] CHAKRABORTY A, JOSHI P K, SACHDEVA K. Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region[J]. Ecological Engineering, 2016, 97: 593
文章摘自:陈思如,张任翔,张学治等.基于MaxEnt优化模型的罗布麻类植物潜在分布研究[J].北京师范大学学报(自然科学版),2023,59(03):377-387.
