摘 要:高温干旱是影响作物生长及最终生产力的主要胁迫源。当前,无人机遥感技术已在作物倒伏和病虫害的分级监测研究中取得重大进展,但有关利用无人机遥感进行作物抗旱等级监测的研究却鲜有报道。因此,以苎麻种质资源为研究对象,提出了苎麻抗旱性量化标准,并提供了一种利用无人机多光谱遥感鉴定苎麻种质资源抗旱性的方法。首先,由专家对36份苎麻种质资源进行抗旱性分级;然后,结合无人机多光谱遥感获取的植被指数,采用随机森林(Random forest,RF)、支持向量机(Support vector machine,SVM))、决策树((Decision tree,DT)3种机器学习方法分别构建苎麻抗旱性鉴定模型,并通过苎麻在高温干旱胁迫下的表型响应检验鉴定结果;最后,基于无人机获取的遥感表型,筛选高温干旱胁迫下优质苎麻种质资源。结果表明,利用SVM构建的苎麻抗旱性鉴定模型正确率达到0.74,不同抗旱级分类F1得分范围为0.69~0.79,说明该方法能用于苎麻种质资源抗旱性评估。利用无人机遥感数据反演得到的3项苎麻表型性状(叶绿素相对含量、叶面积指数、株高)均与人工测量值具有较强的相关性,在此基础上,研究从高温干旱胁迫中筛选出了3个优质苎麻种质资源PJ-CD、WS-XM、湘苎7号。
关键词:高温干旱胁迫;苎麻;抗性筛选;无人机遥感
0 引言
高温和干旱是影响作物生产的重要胁迫源。随着全球气候变化,高温和干旱在未来很可能更加频繁地重合发生,导致局部地区作物安全受到威胁[1-2]。作为重要的纤维作物,苎麻是最易受高温干旱胁迫影响的作物之一[3],在高温干旱胁迫下,苎麻光合作用受到影响,产量和纤维质量将显著降低[4]。因此,迫切需要挖掘苎麻种质资源,快速筛选出适应未来气候变化的抗旱苎麻品种。
作物抗旱性的量化是解决这一问题的关键。高温干旱胁迫会引起作物一系列形态和生理变化[5],因此利用表型指标反映作物抗旱性的方法被提出(间接量化)。在这种方法中,育种家和研究人员将高温干旱胁迫下作物的表型响应作为抗旱性的量化指标[6],进而缓慢地筛选并培育更高抗性的品种。其中,高温干旱胁迫下的最终生产力是评估作物耐旱性最有效、直观的表型[7]。产量组成因素(如株高、分株力、结实率)[8]、叶面积[9]、含水率、叶片颜色[10]、蒸腾速率[11-12]等也是常用的评估作物抗旱性的重要表型指标,如文献[13]根据块茎淀粉产量评估了马铃薯抗旱性,并利用汽车激光雷达系统揭示了与耐旱性相关的表型(株高、叶面积、投影叶面积、叶角)差异。文献[14]利用高通量方法优化了高温干旱胁迫下小麦表型监测。以上研究都是采用间接量化方法评估作物抗旱性,存在以下弊端。第一,过于依赖表型数据,但表型却受基因型和环境因素的复杂交互影响[15],是高温干旱与其他非生物胁迫源相互作用下呈现的外部反映。第二,作为抗旱性量化的表型指标通常采用随机采样方式获取,无法反映品种的整体情况,缺乏跟踪个体的生长轨迹。
直接量化是根据农艺学评估标准,直接对作物种质资源抗性进行分级量化的一种方法。这种分级量化方法已经在作物倒伏程度分类[16-17]、病虫害程度监测[18-22]中得到了广泛应用,但是有关作物抗旱性量化监测的研究还鲜有报道,基于无人机多光谱遥感评价苎麻抗旱性的研究更是少见。因此,本文建立苎麻种质资源抗旱性评估标准,构建基于无人机多光谱遥感的苎麻种质资源抗旱性鉴定模型,研究高温干旱胁迫下苎麻表型的响应,以及苎麻生长与基因型、环境的关系,以实现高温干旱胁迫下基于遥感表型的优质苎麻种质资源筛选。
1 材料与方法
1.1 试验设计与环境条件
高温干旱试验于2022年7-8月湖南省长沙市湖南农业大学苎麻基地(28°11¢01"N,113°04¢10"E)进行。试验期间,该地区持续晴朗,未降雨,维持了旱情。在进行了持续55d高温干旱胁迫处理后,于2020年8月25日对苎麻种质资源抗旱性进行评估。供试材料由36份苎麻种质资源组成,设2次重复,共72个小区。每小区尺寸为2.0m×1.8m,3行×4蔸,蔸间距0.4m,行间距0.6m,排水沟宽0.5m,每个小区面积约为12m2(图1)。
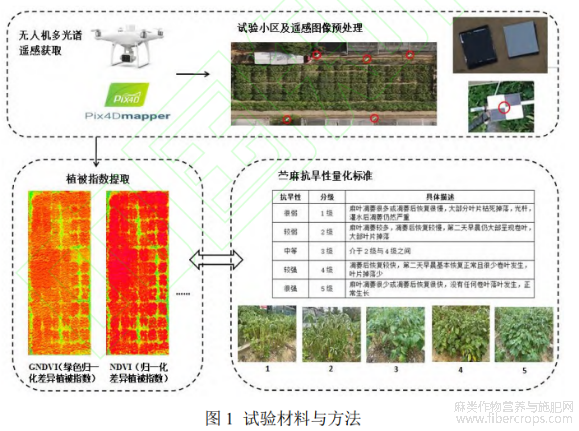
采用气象观测系统持续记录气象要素,包括光照强度、大气压强、大气温度、大气湿度、降水量等,数据每隔1h记录1次。如图2所示,2022年7月来,该地日最高温度多在35℃以上,平均最高温度达到36.5℃,最低温集中于27~30℃,最低温平均值为27.05℃。试验期间每日累积光照强度范围集中于500000~600000Lux,日平均光照为503319Lux,无有效降雨天数,日平均降水量仅为2.33mm,平均大气相对湿度为75.99%。
![]()
1.2 无人机多光谱数据采集与预处理
采用大疆精灵4pro无人机收集多光谱数据,该无人机遥感系统采用RTK厘米级定位系统,集成了6个相机(包括红光、绿光、蓝光、RGB、近红外、红边),每个相机有效像素为200万(图1)。执行飞行任务时,无人机飞行高度为30m,云台俯仰角为-90°,设置主航向上重叠率80%,主航线间图像重叠率70%。8月25日当天,于09:30、11:15、15:45采集3次遥感数据。
使用Pix4Dmapper软件对3次飞行的影像进行拼接,生成试验区域的数字正射影像和数字表面模型。根据反射板实际反射率对影像进行辐射校正,为获取准确的地理参考,5个地面控制点的坐标位置被引入用于地理校正(图1)。采用PhenoAI air软件提取每个苎麻小区的多光谱数据,PhenoAI air能够对遥感影像进行自动化去噪和背景分割,并根据计算出苎麻植被部分的植被指数(Vegetation index,VIs)(图1)。为避免沟壑以及作物生长的边缘效应影响,在每个小区内部建立感兴趣区域(Regions of interest,ROI),并将每个小区分割为4个ROI以扩充数据集的大小。在小区抗旱性表现均一的情况下,这种扩充方法能够提供更加丰富的样本材料,最终总数据集由864个样本组成。
1.3田间数据采集
根据苎麻种质资源抗旱性分级标准[23],5个抗旱等级被设计出来,包括抗旱性很强、强、中等、弱、很弱(图1)。在苎麻接受持续高温干旱处理后,由湖南农业大学苎麻生理研究专家进行官方诊断,并综合多名专家意见对36份苎麻种质资源进行最终的抗旱性评估。评估结果表明,不同抗旱性苎麻分布较为均匀,其中1、2级抗旱品种均7个,占比19.40%;3级抗旱品种8个,占比22.20%;4级抗旱品种9个(25%);5级抗旱品种5个,占比13.90%,36个苎麻品种抗旱性的变异系数为1.8,苎麻群体的抗旱性存在显著的遗传变异。
于2022年7月15日、8月1日、8月25日人工测量苎麻种质资源生长指标,包括株高、叶面积指数、叶绿素相对含量(SPAD值)。株高利用直尺测量苎麻植株底部至冠层顶端的距离;叶面积指数(Leaf area index,LAI)采用LAI-2200型冠层分析仪测量;SPAD值通过日本KONICAMINOLTA公司生产的SPAD-502型叶绿素仪测量,以上数据均通过随机选取各小区10株苎麻求取平均值获取。
1.4 相关性分析、ANOVA方差分析
苎麻抗旱等级为非连续变量,因此采用肯德尔相关系数进行相关性分析。肯德尔相关系数是一种秩相关系数,所计算的对象是分类变量。研究计算了42项VIs与苎麻抗旱性的相关性,VIs与抗旱性之间具有的强相关是实现遥感鉴定苎麻抗旱性的前提。
采用ANOVA分析进一步确定抗旱苎麻品种与高温干旱敏感品种的光谱差异。ANOVA分析允许对多样本进行两两比较,分析不同抗旱级组内变异对总变异的贡献,从而筛选出能够反映不同抗旱级苎麻高温干旱响应的光谱描述符。相关性分析和ANOVA分析均在SPSS中进行,分析结果采用Origin进行可视化。
1.5 模型构建与评估
以遥感获取的VIs为自变量,实测苎麻抗旱等级为因变量,采用随机森林(Random forest,RF)、支持向量机(Support vector machine,SVM)、决策树(Decision tree,DT)3种机器学习方法构建苎麻抗旱性鉴定模型。以遥感获取的VIs为自变量,实测苎麻表型数据为因变量,采用偏最小二乘回归(Partial least squares regression,PLSR)方法构建苎麻表型监测模型,进而从遥感影像中得到估测的SPAD值(SPAD_UAV)、估测的叶面积指数(LAI_UAV)和估测的株高(PH_UAV)。对于每种模型,按照7:3的比例随机划分训练集和验证集,通过网格搜索和五折交叉验证在训练集中确定各算法的最佳模型参数,然后分别以最佳参数进行训练,将训练好的各算法在验证集中评估模型表现。
精确率、召回率、F1得分、正确率4项指标被用于评估苎麻抗旱性鉴定模型在训练和测试集上的性能。精确率反映正确预测样本占所有预测样本的比例,召回率表示在所有正样本中检出为正样本的数量,F1得分是精确率和召回率的加权调和平均,正确率表示预测正确的占所有数量的比例。以上指标采用“classification_report”计算。R2和均方根误差(Root mean square error,RMSE)被用于评价苎麻表型监测模型的性能,R2越高,RMSE越低,表明模型拟合精度越高越稳定。
2 结果与分析
2.1 不同抗旱等级苎麻的VIs差异
为探讨利用多光谱信息鉴定苎麻抗旱性的可行性,对苎麻抗旱性和VIs进行相关性分析。如图3a所示,09:30、11:30、15:453个时间点的VIs与苎麻抗旱性的相关性均呈显著或极显著水平,肯德尔系数在-0.40~0.50之间。其中,RVI(比值植被指数)、MSR(改进型比值植被指数)、WDRVI(宽动态范围植被指数)与苎麻抗旱性呈极显著正相关,3个时期的累计肯德尔系数分别达到1.077、1.049、1.021。GRNDVI(绿红归一化植被指数)、RGRI(红绿比值指数)、R(红光波段反射率)与苎麻抗旱性呈极显著负相关,3个时期的累计肯德尔系数分别为-0.856、-0.834、-0.766。不同时间点获取的VIs与抗旱性的相关性表现出逐渐低的趋势,09:30获取的VIs与苎麻抗旱性达到最大相关性,相关性系数为11.887,比后两个时间段分别高出14.684%、15.937%。
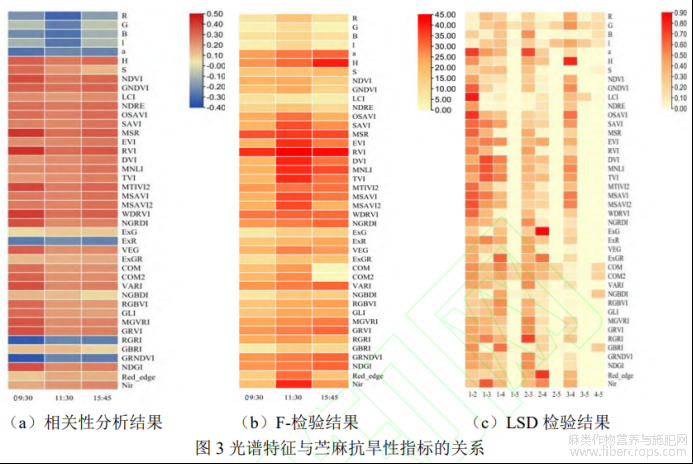
采用ANOVA方法分析不同抗旱等级苎麻的VIs差异。由F-检验结果可知(图3b),3个时间点的平均F值分别为18.893、26.249、21.931,不同抗旱级苎麻的VIs存在显著组间差异,VIs具有区分苎麻抗旱性的能力。计算LSD值以进一步确定不同抗旱级苎麻组内存在的差异(图3c),5级与其他抗旱等级的平均LSD均低于0.05,这表明抗旱性强的苎麻种质资源具有独特的光谱特征,与其他抗旱等级苎麻在VIs上存在显著差异。NDRE(归一化差异红色边缘指数)、NGBDI(归一化绿蓝差异指数)和S(饱和度)在多重比较中具有最低的LSD,说明这3个参数能够作为苎麻水分亏缺和高温反应的光谱描述符,用于区分不同苎麻抗旱等级。
2.2 苎麻抗旱性鉴定模型
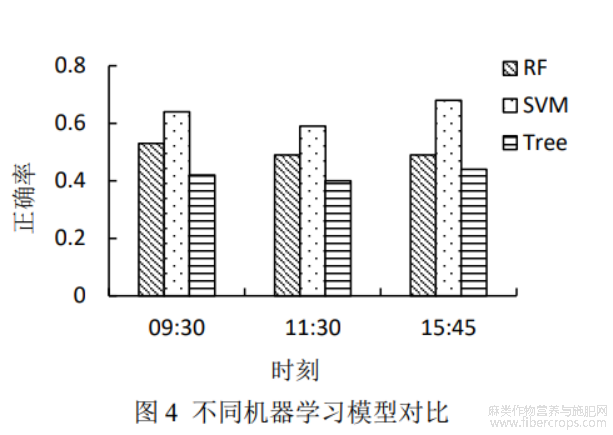
结合计算得到的42个VIs,采用机器学习方法构建苎麻抗旱性鉴定模型。图4对比了3种机器学习方法的鉴定性能。由图4可知,3种模型都具有较高的精度,RF、SVM、DT的正确率范围分别为0.42~0.64、0.40~0.59、0.44~0.68。其中,SVM模型具有最佳正确率,3个时间点的平均正确率达到0.64,其次为RF(0.50)、DT(0.42)。从监测时间看,模型精度从大到小整体表现依次为15:45、09:30、11:30。随着白天温度逐渐升高,蒸发量增强,不同抗旱级苎麻的表型差异加大,在白天晚些时间,差异达到峰值,因此下午的监测结果比上午、中午更好。高温干旱胁迫后的恢复速度及程度也是苎麻抗旱性的重要评价标准,由于夜晚的水分处理及温度的降低,抗旱性更强的品种恢复更好,这种差异在第2天上午得到反映。
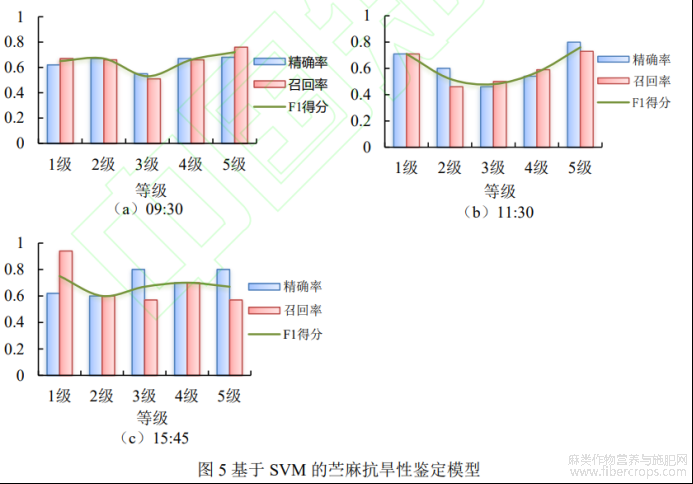
对于每个抗旱级,采用SVM模型分析测试集的鉴定结果(图5)。09:30时,模型正确率为0.64,不同抗旱级分类精度范围为0.55~0.68,召回率范围为0.51~0.76,F1得分范围为0.53~0.72。11:15时,模型正确率为0.59,不同抗旱级分类精度范围为0.46~0.80,召回率范围为0.46~0.73,F1得分范围为0.48~0.76。15:45时,模型正确率为0.68,不同抗旱级分类精度范围为0.60~0.80,召回率范围为0.57~0.94,F1得分范围为0.60~0.75。F1得分呈现“两级高,中间低”的趋势,这表明相比抗旱性中等的苎麻,抗旱极强和极弱的苎麻更容易识别,这与苎麻不同抗旱等级间的变异分析结果一致。
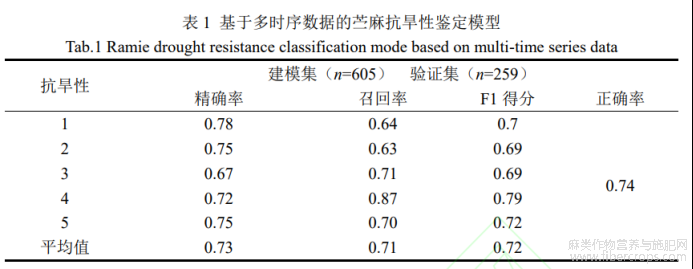
结合不同时间段获取的遥感数据有可能提高最终分类准确率,因此研究以3个时间点获取的遥感参数为自变量,苎麻种质资源抗旱级别为因变量,采用SVM构建了基于多时序的苎麻抗旱性鉴定模型(表1)。由表1可知,多时序模型正确率达到0.74,比单一时期模型评价正确率提升16.23%。不同抗旱级分类精度范围为0.67~0.78,平均精度为0.73,召回率范围为0.63~0.87,平均召回率为0.71,F1得分范围为0.69~0.79,平均值为0.72。
2.3 基于无人机遥感的苎麻表型监测结果
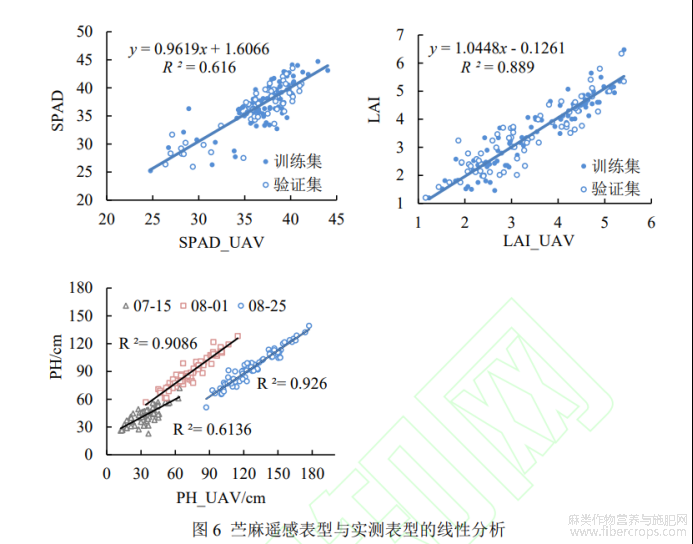
采用PLSR构建苎麻表型监测模型(表2),SPAD值估测模型和LAI估测模型在训练集和验证集都得到了较为理想的监测效果。对于苎麻SPAD值,训练集和验证集的R2分别为0.648、0.756,RMSE分别为2.362、2.227;相比SPAD值,多光谱遥感对LAI的监测精度更高,训练集和验证集的R2分别为0.885、0.778,RMSE分别为0.396、0.519。
图6为苎麻遥感表型与实测表型的线性分析。线性相关分析结果表明,遥感估测的SPAD_UAV、LAI_UAV、PH_UAV与人工测量值具有较强的相关性,SPAD_UAV与SPAD的R2为0.616,LAI_UAV与LAI的R2为0.889。从数字表面模型(Digital surface model,DSM)计算了不同生育期的PH_UAV,不同生育期PH_UAV与PH的拟合效果不同,随着生育期的后移,PH_UAV和PH的R2逐渐提高,到8月25日达到0.926。这是因为,早期DSM混合了裸露土表,导致混合像元低于实际值。较理想的表型监测结果表明,无人机遥感是快速、无损、精准监测环境胁迫下作物表型的工具,监测结果可用于高温干旱胁迫下苎麻特异表型的分析。

2.4 高温干旱胁迫下苎麻表型响应
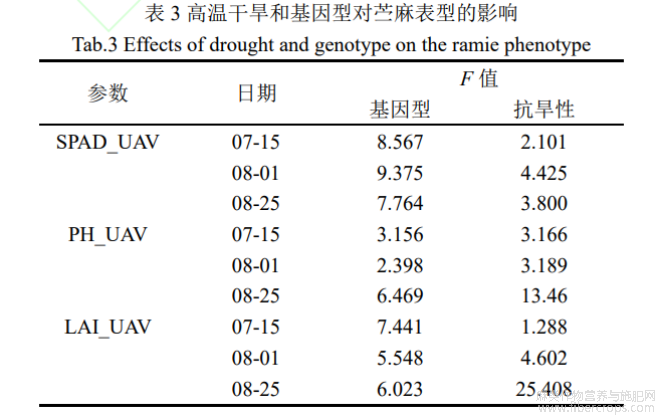
利用ANOVA方法分析高温干旱胁迫下苎麻不同生育期的表型差异(表3)。不同苎麻种质资源在表型上存在显著差异,F值范围分别达到8.567~9.375、2.398~6.469、5.548~7.441。而高温干旱胁迫使得品种间的表型差异更为明显,表现为不同抗旱性苎麻具有较为明显的组间差异,F值在PH_UAV和LAI_UAV上有所提高。随着时间的后移,不同抗旱性苎麻种质资源的表型差异逐渐增大,这一表现在8月25日测量得到的PH_UAV和LAI_UAV上更为突出,F值分别为13.460、25.408,表明PH_UAV和LAI_UAV苎麻对高温干旱十分敏感。
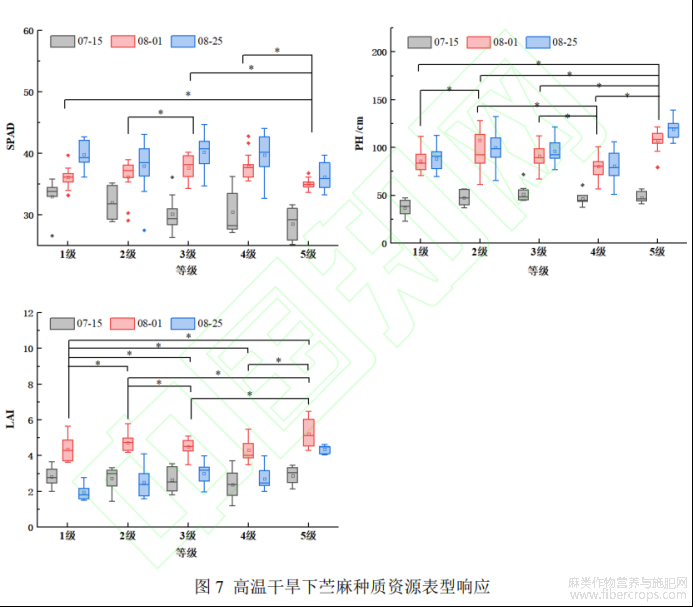
采用LSD进一步分析8月25日不同抗旱级苎麻表型的组内差异(图7,图中*表示具有显著差异)。2级与3级、5级与1、3、4级在SPAD_UAV上存在显著差异;高温干旱对苎麻出苗及生长速度有影响,在苗期,随着苎麻抗旱能力的增强,苎麻生长速度越快,株高越高。到苎麻生长后期,1级与2级、4级与2、3级、5级与1、2、3、4级在PH_UAV上均呈现显著差异;随着时间的后移,LAI_UAV先增大后减少,在其生育后期除4级与2、3级之间没有显著差异外,各不同抗旱级苎麻的LAI_UAV均存在显著差异。这是因为,为应对高温干旱胁迫,苎麻叶片会逐渐脱落并且卷曲,减少由于叶面积过大导致的强蒸腾作用。PH_UAV和LAI_UAV在不同抗旱级的差异表现,说明这两个生长参数能够成为耐旱标志。
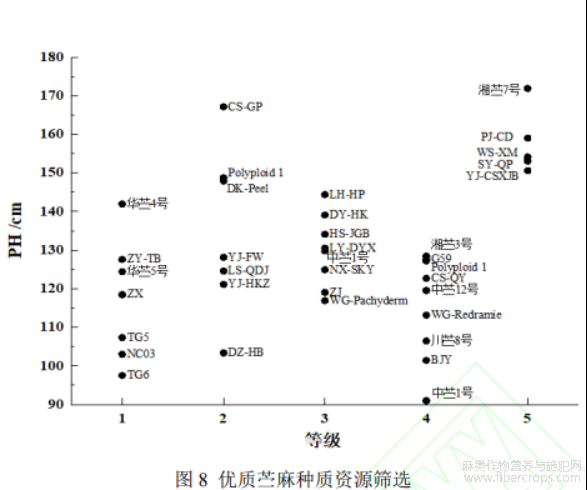
2.5 高温干旱胁迫下优质苎麻种质资源筛选
通过分析不同抗旱等级苎麻的遥感表型,能够根据特定的生产要求筛选优质苎麻种质资源。不同抗旱级苎麻遥感表型信息如图8,图中字母及文字表示苎麻品种。结果表明,在具有5级抗旱能力的苎麻种质资源中,PJ-CD、WS-XM、湘苎7号在株高和叶面积上都有较好的表现。株高和叶面积是评估苎麻产量的重要指标,综合这两项指标可以筛选PJ-CD、WS-XM、湘苎7号为高温干旱胁迫下高产苎麻品种。在文献[24]等的研究中,WS-XM被鉴定为高温干旱期高产优质的苎麻饲用种质资源,文献[25]等的研究表明在持续干旱条件下,湘苎7号表现出较强的生长活性,以上研究结果验证了筛选结果。另外,PJ-CD的发现也能为高温干旱胁迫下,苎麻品种选育提高参考。

3 结论
(1)不同抗旱等级苎麻表现出的光谱数据存在显著组间差异,其中,抗旱性很强的苎麻种质资源(5级)与其他抗旱等级苎麻在VIs上具有最大差异,表明遥感提取的VIs具有区分苎麻抗旱性的能力。NDRE、NGBDI和S这3个光谱参数在多重比较中具有最低的LSD,能够作为苎麻水分亏缺和高温反应的光谱描述符。
(2)分别采用RF、SVM、DT3种机器学习算法构建苎麻抗旱性鉴定模型,结果表明3种模型都具有较高的精度,正确率范围分别为0.42~0.64、0.40~0.59、0.44~0.68。其中,基于多时序数据的SVM模型具有最佳正确率(0.74),F1得分呈现“两级高,中间低”的趋势,表明抗旱极强和极弱的苎麻更容易识别。
(3)利用无人机遥感数据估测得到的SPAD_UAV、LAI_UAV、PH_UAV与人工测量值具有较强的相关性,可用于监测高温干旱胁迫下不同抗旱级苎麻表型差异,其中SPAD_UAV与SPAD的R2为0.616,LAI_UAV与LAI的R2为0.889,PH_UAV和PH的R2为0.614~0.926。结果表明,高温干旱胁迫使得品种间的表型差异更为明显,抗旱苎麻品种出苗更快、长得更高、叶面积更高。
参考文献
[1] BASU S, RAMEGOWDA V, KUMAR A, et al. Plant adaptation to drought stress [J]. F1000Research, 2016, 5:1554.
[2] HARRISON M T, TARDIEU F, DONG Z S, et al. Characterizing drought stress and trait influence on maize yield under current and future conditions [J]. Global Change Biology, 2014, 20:867-878.
[3] 刘飞虎, 张寿文, 梁雪妮, 等. 干旱胁迫下不同苎麻品种的形态解剖特征研究[J]. 中国麻作, 1999 (4):2-7.
LIU Feihu, ZHANG Shouwen, LIANG Xueni, et al. Morphological and anatomical characteristics of different ramie varieties under drought stress[J]. Plant Fiber Sciences in China, 1999(4):2-7.
[4] 王继龙, 刘婕仪, 刘皖慧, 等. 苎麻响应水分胁迫研究进展[J]. 中国麻业科学, 2020, 42(4):178-186.
WANG Jilong, LIU Jieyi, LIU Wanhui, et al. Research progress in response to water stress in ramie[J]. Plant Fiber Sciences in China, 2020, 42(4):178-186.
[5] COSTA J M, GRANT O M, CHAVES M M. Thermography to explore plant environment interactions[J]. Journal of Experimental Botany, 2013, 64:3937-3949.
[6] ALTAMIMI N, LANGAN P, BERNÁD V, et al. Capturing crop adaptation to abiotic stress using image-based technologies[J]. Open Biol., 2022, 12:210353.
[7] RICHARDS R A, HUNT J R, KIRKEGAARD J A, et al. Yield improvement and adaptation of wheat to water-limited environments in Australia-a case study [J]. Crop Pasture Sci., 2014, 65:676-689.
[8] ALICHE E B, OORTWIJN M, THEEUWEN TPJM, et al. Drought response in field grown potatoes and the interactions between canopy growth and yield [J]. Agric Water Manag, 2018,206:20-30.
[9] WELLSTEIN C, POSCHLOD P, GOHLKE A, et al. Effects of extreme drought on specific leaf area of grassland species: a meta-analysis of experimental studies in temperate and sub-Mediterranean systems[J]. Global Change Biology, 2017, 23(6): 2473-2481.
[10] FU F J, ZHAO Z, ZHANG W X. Study on photosynthetic indexes and drought resistance of maize under water stress[J]. Journal of Mountain Agriculture and Biology, 2004, 23(6): 471-474.
[11] ELHABTI A, FLEURY D, JEWELL N, et al. Tolerance of combined drought and heat stress is associated with transpiration maintenance and water soluble carbohydrates in wheat grains [J]. Frontiers in Plant Science, 2020, 11:1555.
[12] HAAS M, SPRENGER H, ZUTHER E, et al. Can metabolite and transcript-based selection for drought tolerance in Solanum tuberosum replace selection on yield in arid environments?[J]. Frontiers Plant Sci., 2020, 11:1071-1087.
[13] MULUGETA A G, HAAS M, Köhl K. LIDAR-based phenotyping for drought response and drought tolerance in potato[J]. Potato Res., 2022.
[14] CORREIA P, CAIRO W J, BERNARDES D S A, et al. High-throughput phenotyping of physiological traits for wheat resilience to high temperature and drought stress[J]. J. Exp. Bot., 2022, 73(15):5235-5251.
[15] 周济, TARDIEU F, PRIDMORE T, 等. 植物表型组学:发展、现状与挑战[J].南京农业大 学学报, 2018,41(4) : 580-588.
ZHAO Ji, TARDIEU F, PRIDMORE T, et al. Plant phenoomics: development, current situation, and challenges[J]. Journal of Nanjing Agricultural University, 2018,41(4) : 580-588.
[16] YANG X, GAO S, SUN Q, et al. Classification of maize lodging extents using deep learning algorithms by UAV-based RGB and multispectral images[J]. Agriculture, 2022, 12(7): 970.
[17] 章凌翔. 基于无人机多光谱影像油菜产量预测及倒伏风险评价[D].西北农林科技大 学,2022.
[18] 任志鹏, 高睿, 王大庆. 基于哨兵 2 号多光谱影像的水稻倒伏识别与分类[J].节水灌溉, 2022(7):44-50.
REN Zhipeng, GAO Rui, WANG Daqin. Rice lodging recognition and classification based on Sentinel-2 multispectral image[J]. Water Saving Irrigation, 2022(7):44-50.
[19] AHMAD A, SARASWAT D, EL GAMAL A. A Survey on using deep learning techniques for plant disease diagnosis and recommendations for development of appropriate tools[J]. Smart Agricultural Technology, 2022, 3: 100083.
[20] GHOSAL S, BLYSTONE D, SINGH AK, et al. An explainable deep machine vision framework for plant stress phenotyping[J]. Proc. Natl. Acad. Sci. USA., 2018, 115(18):4613-4618.
[21] 姜红花, 杨祥海, 丁睿柔, 等. 基于改进 ResNet18 的苹果叶部病害多分类算法研究 [J/OL]. 农业机械学报:1-9.
JIANG Honghua, YANG Xianghai, DING Ruirou, et al. Research on multi-classification algorithm of apple leaf diseases based on improved ResNet18[J/OL]. Transactions of the Chinese Society for Agricultural Machinery: 1-9.
[22] 李凯雨, 朱昕怡, 马浚诚, 等. 基于混合扩张卷积和注意力的黄瓜病害严重度估算方法 [J/OL].农业机械学报:1-12.
LI Kaiyu, ZHU Xinyi, MA Juncheng, et al. Estimating cucumber disease severity based on mixed expanded convolution and attention[J/OL]. Transactions of the Chinese Society for Agricultural Machinery:1-12.
[23] 粟建光,戴志刚. 中国麻类作物种质资源及其主要性状[M]. 北京:中国农业出版社,2016.
[24] 刘俊欢, 揭雨成, 邢虎成, 等. 高温干旱期 203 份苎麻饲用种质资源的农艺性状鉴定评 价及高产优质种质筛选[J]. 中国农学通报, 2016, 32(3):54-60.
LIU Junhuan, JIE Yucheng, XIN Hucheng, et al. Identification and evaluation of agronomic characters of 203 ramie feeding germplasm resources and screening of high yield and high quality germplasm during high temperature and drought[J]. Chinese Agricultural Science Bulletin, 2016, 32(3):54-60.
[25] 李林林, 马渊博, 张浪, 等. 3个苎麻品种对干旱胁迫的生理生化响应[J]. 中国麻业科学, 2018, 40(1):22-27. LI Linlin, MA Yuanbo, ZHANG Lang, et al. Physiological and biochemical responses of three ramie varieties to drought stress[J]. Plant Fiber Sciences in China, 2018, 40(1):22-27.
文献摘自:[1]付虹雨,王薇,卢建宁,岳云开,崔国贤,佘玮.基于无人机多光谱的耐旱苎麻品种筛选[J/OL].农业机械学报:1-15[2023-03-15].http://kns.cnki.net/kcms/detail/11.1964.S.20230221.1453.040.html
