摘 要:试验旨在研究不同膨化亚麻籽日粮饲喂程序对生长育肥猪生长性能、屠宰性能、脂肪酸沉积及免疫功能的影响。选取初始体重为(36.76±0.49)kg的杜×巴×嘉生长育肥猪120头,随机分为4组,每组3个重复,每个重复10头,试验期90d。对照组全程饲喂基础日粮,T30、T60和T90组先饲喂基础日粮并分别于屠宰前30、60、90d开始饲喂5%膨化亚麻籽日粮。结果表明:与对照组相比,不同膨化亚麻籽日粮饲喂程序对生长性能和屠宰性能均无显著影响,但均提高了背最长肌和背膘ω-3多不饱和脂肪酸(ω-3PUFA)含量、降低了ω-6/ω-3PUFA比值(P<0.05),且改善程度随着饲喂时间的增加而增加(P<0.05);各组间总蛋白(TP)、白蛋白(ALB)、谷丙转氨酶(ALT)、谷草转氨酶(AST)和碱性磷酸酶(ALP)等血清生化指标均无显著差异;与对照组相比,T30和T60组血清补体3(C3)含量均升高(P<0.05),T60组血清补体4(C4)、免疫球蛋白M(IgM)和免疫球蛋白G(IgG)含量均升高(P<0.05)。综上,屠宰前90d开始饲喂添加5%膨化亚麻籽的日粮不影响生长肥育猪生长性能、屠宰性能和机体免疫功能,并可显著提高背最长肌和背膘ω-3PUFA含量,ω-6/ω-3PUFA比值可达到中国营养学会推荐范围。
关键词:生长育肥猪;膨化亚麻籽;饲喂程序;生长性能;脂肪酸沉积
我国是世界最大的猪肉生产国和消费国,当前国内生猪养殖规模已可满足本国市场需求,人们对猪肉的消费需求已经从数量转为品质,猪肉中高端消费分级市场逐渐形成。中高端猪肉的生产途径主要包括优良猪种开发(如中国地方猪及其配套系)、营养调控和饲养管控,营养调控是优化猪肉脂肪酸组成的有效途径之一。猪肉脂肪中的脂肪酸组成是评价猪肉营养效价的重要指标。猪肉中的多不饱和脂肪酸(Polyunsaturated Fatty Acids,PUFA)主要分为ω-3多不饱和脂肪酸(ω-3Polyunsaturated Fatty Acids,ω-3PUFA)和ω-6多不饱和脂肪酸(ω-6Polyunsaturated Fatty Acids,ω-6PUFA)。Ω-3PUFA在预防心血管疾病、维系大脑生长发育以及调节机体免疫力等方面具有重要作用[1-4];而过高的ω-6/ω-3PUFA比值能引起心血管疾病、糖尿病和自身免疫性疾病等很多疾病[5-10]。在生产实践中,提高猪肉产品中ω-3PUFA含量、降低ω-6/ω-3PUFA比值有助于提升猪肉营养价值与品质。
亚麻籽(油)中的ω-3PUFA主要是α-亚麻酸(α-Linolenic Acid,ALA),是自然界中ALA含量最丰富的植物性原料,ALA约占其脂肪酸总量的50%以上[11]。大量研究表明,日粮中添加亚麻籽(油)可以提高猪肉中ω-3PUFA含量,并可降低ω-6PUFA含量,从而降低ω-6/ω-3PUFA比值[11-13]。因此,本试验旨在研究不同膨化亚麻籽日粮饲喂程序对生长育肥猪生长性能、屠宰性能、脂肪酸沉积及免疫功能的影响,以期为富ω-3PUFA猪肉的生产实践提供理论依据。
1材料与方法
1.1试验材料
杜×巴×嘉生长育肥猪(杜洛克×巴克夏×嘉兴黑猪)由嘉兴市青莲养殖科技有限公司猪场提供,并在该猪场开展试验。膨化亚麻籽由嘉兴红太阳饲料有限公司提供(粗蛋白质含量为21.0%、粗脂肪含量为55.0%、ALA含量为19.7%)。试验日粮分为基础日粮和膨化亚麻籽日粮,参照NRC(2012)猪营养需要,并结合嘉兴黑猪耐粗饲及猪场实际情况配制,委托海盐申港饲料有限公司生产,日粮组成和营养成分见表1。
表1 日粮组成和营养成分(风干基础)
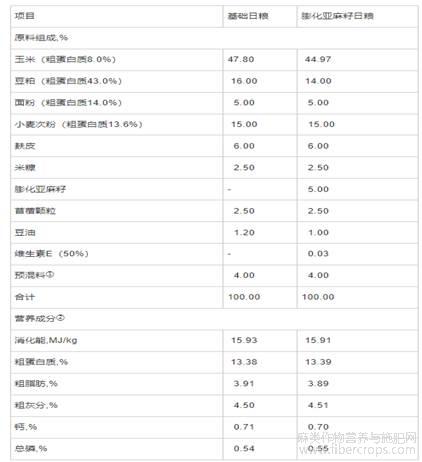
注:①预混料为每千克日粮提供:维生素A8000IU,维生素B14mg,维生素B23.6mg,维生素B540mg,维生素B64mg,维生素B120.02mg,维生素D33000IU,维生素E20IU,维生素K32mg,生物素0.15mg,叶酸1.0mg,D-泛酸11mg,烟酸10mg,抗氧化剂100mg,铜10mg,铁80mg,锰80mg,锌75mg,碘0.40mg,硒0.30mg。②营养成分中消化能为计算值,其余为实测值。
1.2试验设计与饲养管理
采用单因素完全随机设计,选取120头初始体重(36.76±0.49)kg的健康杜×巴×嘉生长育肥猪,公母各半,随机分为4组,每组3个重复,每个重复10头猪。试验期为90d。对照组全程饲喂基础日粮,T30、T60和T90组先饲喂基础日粮并分别于屠宰前30、60、90d开始饲喂膨化亚麻籽日粮。试验期间自由采食和饮水,保持圈舍清洁,消毒和防疫等其他管理按猪场常规程序进行。
1.3样品采集与处理
试验结束后,将各组试验猪全部运送至浙江青莲食品股份有限公司新兴屠宰场,经24h停食静养后进行屠宰,参照《瘦肉型猪胴体性状测定技术规范》(NY/T825-2004)测定宰前活重和胴体重。经麻电击晕后,用真空促凝管采血10mL,室温下倾斜放置30min,3000r/min离心15min,吸取上清并分装于1.5mL离心管中,-20℃保存。胴体经劈半后,取左半侧测定背膘厚、眼肌长和宽。取左半侧胴体背最长肌和背膘各50g,包装并编号,-20℃冻存备用。
1.4测定指标与方法
1.4.1生长性能与屠宰性能
分别于试验开始和试验结束当天08:00空腹称重,通过自动饲喂系统采集记录试验过程中各组每天的采食量,计算平均日增重(ADG)、平均日采食量(ADFI)和耗料增重比(F/G)。屠宰性能计算公式:
屠宰率=胴体重/宰前活重×100%
眼肌面积=眼肌长×眼肌宽×0.7
1.4.2脂肪酸组成测定
脂肪酸组成按照《食品安全国家标准食品中脂肪酸的测定》(GB5009.168-2016)测定。各类脂肪酸含量用其占总脂肪酸质量百分比的形式表示,并根据各类脂肪酸的占比计算饱和脂肪酸(Saturated Fatty Acids,SFA)含量、PUFA含量、ω-3PUFA含量、ω-6PUFA含量和ω-6/ω-3PUFA比值。计算公式:
SFA=丁酸(C4:0)+己酸(C6:0)+辛酸(C8:0)+葵酸(C12:0)+十一碳酸(C11:0)+十二碳酸(C12:0)+十三碳酸(C13:0)+豆蔻酸(C14:0)+十五碳酸(C15:0)+棕榈酸(C16:0)+十七烷酸(C17:0)+硬脂酸(C18:0)+花生酸(C20:0)+二十一碳酸(C21:0)+山嵛酸(C22:0)+二十三碳酸(C23:0)+木焦油酸(C24:0)
PUFA=反,反-9,12-十八碳二烯酸(C18:2n6t)+亚油酸(C18:2n6c)+顺,顺,顺-6,9,12-十八碳三烯酸(C18:3n6)+亚麻酸(C18:3n3)+花生二烯酸(C20:2)+顺,顺,顺-8,11,14-二十碳三烯酸(C20:3n6)+顺-5,8,11,14-二十碳四烯酸(C20:4n6)+顺-11,14,17-二十碳三烯酸(C20:3n3)+顺-5,8,11,14,17-二十碳五烯酸(C20:5n3)+顺-13,16-二十二碳二烯酸(C22:2n6)+顺-4,7,10,13,16,19-二十二碳六烯酸(C22:6n3)
ω-3PUFA=亚麻酸(C18:3n3)+顺-11,14,17-二十碳三烯酸(C20:3n3)+顺-5,8,11,14,17-二十碳五烯酸(C20:5n3)+顺-4,7,10,13,16,19-二十二碳六烯酸(C22:6n3)
ω-6PUFA=反,反-9,12-十八碳二烯酸(C18:2n6t)+亚油酸(C18:2n6c)+顺,顺,顺-6,9,12-十八碳三烯酸(C18:3n6)+顺,顺,顺-8,11,14-二十碳三烯酸(C20:3n6)+顺-5,8,11,14-二十碳四烯酸(C20:4n6)+顺-13,16-二十二碳二烯酸(C22:2n6)
1.4.3血清生化指标和免疫指标
血清样品送至浙江农林大学教学动物医院,采用全自动生化分析仪(IDEXXCatalystOne®)测定血清总蛋白(TP)、白蛋白(ALB)、谷丙转氨酶(ALT)、谷草转氨酶(AST)和碱性磷酸酶(ALP)。采用酶联免疫法测定血清IgA、IgG、IgM、C3和C4,具体操作按照试剂盒(南京建成生物工程研究所有限公司)说明书进行。
1.5统计分析
数据经Excel初步整理后,采用SPSS26.0软件进行单因素方差分析(One-Way ANOVA),若F检验差异显著则进行Duncan´s多重比较,以P<0.05表示差异显著。试验数据用平均值(Mean)±标准差(SD)表示。
2结果
2.1不同饲喂程序对生长育肥猪生长性能的影响
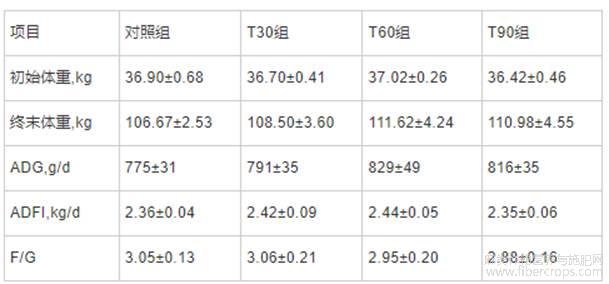
由表2所知,与对照组相比,T30、T60和T90组的生长性能均无显著变化。
表2 不同饲喂程序对生长育肥猪生长性能的影响
2.2不同饲喂程序对生长育肥猪背最长肌脂肪酸组成的影响
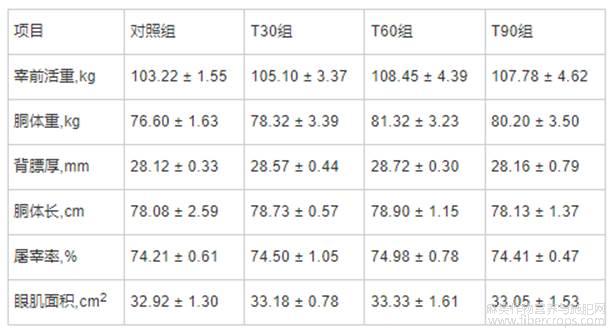
由表3可知,与对照组相比,T30、T60和T90组的宰前活重、胴体重、背膘厚、胴体长、屠宰率和眼肌面积等屠宰性能指标均无显著变化
表3 不同饲喂程序对生长育肥猪屠宰性能的影响
2.3不同饲喂程序对生长育肥猪背最长肌脂肪酸组成的影响
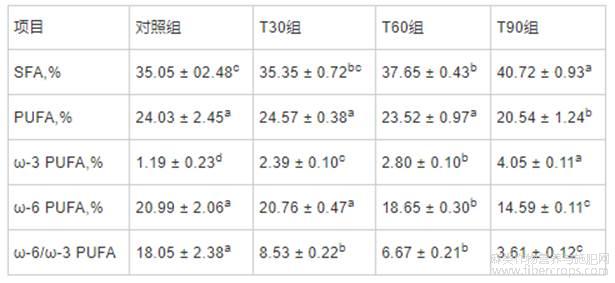
由表4所知,与对照组相比,T90组提高了背最长肌中PUFA含量(P<0.05);与对照组和T30组相比,T60和T90组降低了背最长肌中ω-6PUFA含量(P<0.05),而T60和T90组间无显著差异。在屠宰前饲喂不同时间膨化亚麻籽日粮均可提高背最长肌中ω-3PUFA含量(P<0.05),降低ω-6/ω-3PUFA比值(P<0.05),且随饲喂时间的延长,其作用效果逐渐增强。与对照组相比,T30、T60和T90组背最长肌SFA含量并无显著变化。
表4 不同饲喂程序对生长育肥猪背最长肌脂肪酸组成的影响
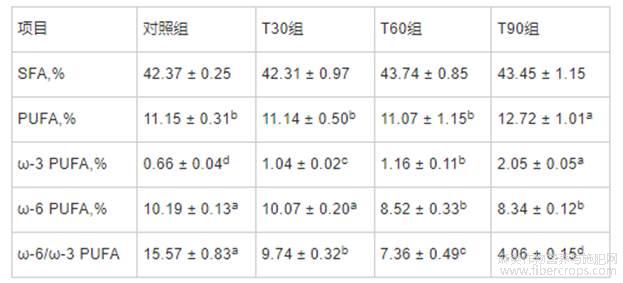
注:同行数据肩标无字母或含相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。下表同。
由表5所知,与对照组相比,各组背膘中SFA含量随着屠宰前膨化亚麻籽日粮饲喂时间的延长而增加,在T90组达到最大值(P<0.05);而T90组背膘中PUFA含量则低于其它组(P<0.05);各组背膘ω-6PUFA含量和ω-6/ω-3PUFA比值随着屠宰前膨化亚麻籽日粮饲喂时间的增加而降低,而ω-3PUFA含量则随着屠宰前膨化亚麻籽日粮饲喂时间的增加而增加,其中T90组ω-3PUFA含量最高,ω-6/ω-3PUFA比值最低(P<0.05)。
表5 不同饲喂程序对生长育肥猪背膘中脂肪酸组成的影响
2.4不同饲喂程序对生长育肥猪血清生化指标和免疫指标的影响
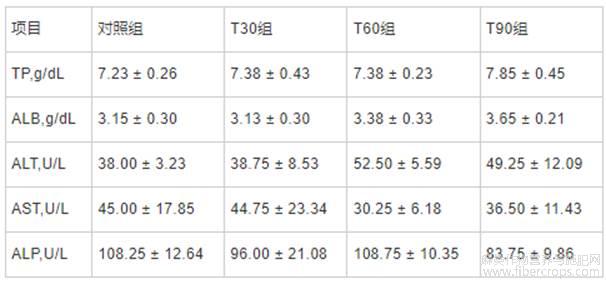
由表6可知,与对照组相比,T30、T60和T90组血清TP、ALB、ALT、AST和ALP等血清生化指标均无显著变化(P>0.05)。
表6 不同饲喂程序对生长育肥猪血清生化指标的影响
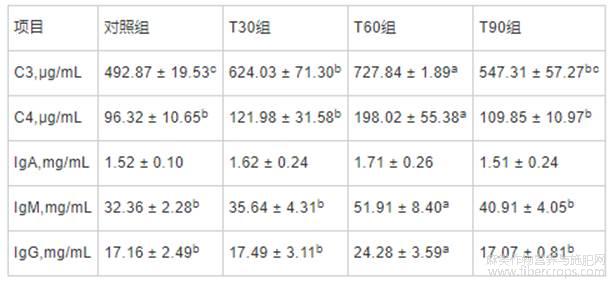
由表7可知,与对照组相比,膨化亚麻籽日粮屠宰前饲喂不同时间未对血清IgA含量产生显著影响。与对照组相比,T30和T60组血清C3含量均升高(P<0.05);T60组血清C4、IgM和IgG含量改善效果显著(P<0.05),其余各组间均无显著差异。
表7 不同饲喂程序对生长育肥猪血清免疫指标的影响
3讨论
3.1不同饲喂程序对生长育肥猪生长性能的影响
亚麻籽是一种优质油料作物,其脂肪含量高且富含ALA。目前,日粮中添加亚麻籽主要用于改善动物产品脂肪酸组成[11,14-17]。大部分研究表明,日粮中添加亚麻籽对猪生长性能无不利影响[18],但各研究中亚麻籽饲喂时间不尽相同。邓波等[13]研究发现,屠宰前40d开始饲喂添加5%和10%亚麻籽日粮对其ADFI和F/G均无显著影响,而在屠宰前72d开始饲喂则显著降低F/G,但对其ADFI无显著性影响。石宝明等[19]研究发现,用含10%亚麻籽的日粮饲喂东北民猪56d,对其生长性能未产生显著影响。本试验以添加5%膨化亚麻籽的日粮分别在屠宰前30、60、90d饲喂生长育肥猪,均未对其生长性能产生影响,较上述研究表现出更长的耐受时间。可能是因为本试验采用的自主培育品种杜×巴×嘉配套系与商品瘦肉型猪种存在一定的生长适应性差异,其带有地方猪血统,具有一定的抗病性和抗逆特性,从而对亚麻籽日粮具有更好的适应性。
3.2不同饲喂程序对生长育肥猪屠宰性能的影响
当前,屠宰企业主要采用白条或分割产品的形式与客户结算。因此,商品猪的屠宰性能可以间接反映各组试验猪的真实商品价值。邓波等[13]研究发现,与对照组相比,分别以添加5%和10%亚麻籽的日粮饲喂生长育肥猪饲喂72d后对其胴体长、屠宰率和背膘厚均无显著影响,与本试验结果基本一致。屠宰性能主要取决于日粮的总体营养水平,而本试验采用等氮等能原则设计日粮,其脂肪酸模式的改变尚不足以引起生长育肥猪的胴体组成变化。Kouba等[20]研究发现,日粮中添加6%亚麻籽饲喂不同时间后(20、60、100d)对其屠宰性能具有显著影响。造成上述差异的原因可能是该试验于同一日龄开始饲喂亚麻籽日粮[20],前者在试验结束时各组间日龄差异较大,而本试验是先饲喂基础日粮并于屠宰前不同时间转换为亚麻籽日粮。
3.3不同饲喂程序对生长育肥猪脂肪酸沉积的影响
作为单胃动物,猪日粮中的脂肪酸经消化道吸收可直接用于合成脂肪而沉积于体内。因此,日粮脂肪酸模式对其脂肪酸组成影响较大,且呈现剂量依赖关系[21-22]。这在本试验结果中也得到印证,即在生长育肥猪日粮中添加5%膨化亚麻籽后,其背最长肌和背膘ω-3PUFA含量均显著增加,且随着屠宰前饲喂时间的增加而增加。Kouba等[20]研究发现,用添加6%亚麻籽的日粮饲喂杜×长×大生长育肥猪20、60、100d均可显著提高其背最长肌和背膘ω-3PUFA含量,却在60d即达到峰值。两者不同的ω-3PUFA沉积效应可能是不同猪种引起的生长速度差异所致。Nuernberg等[23]给生长育肥猪饲喂含5%亚麻籽油日粮至屠宰期(105kg),发现其肌内脂肪和背膘ω-3PUFA含量显著提高,ω-6PUFA含量显著降低。本试验也发现,背最长肌和背膘ω-6PUFA含量随膨化亚麻籽日粮饲喂时间的延长而逐渐降低。可能是由以下2个原因所导致:①由于ALA与亚油酸在合成PUFA时共用去饱和酶和碳链延长酶[1],因此,当ALA含量提高时,会限制亚油酸进一步生成长链ω-6PUFA[23];②油脂在脂肪酶的作用下逐渐水解为游离脂肪酸和甘油,经被动扩散或脂肪酸转运载体进入肠道上皮细胞,之后被脂肪酸结合蛋白质运送至内质网后重新酯化后经肠绒毛进入淋巴系统或门静脉,其中,中链脂肪酸无需脂肪酶的消化可以直接通过门静脉进入肝脏[24],日粮中增加的ω-3PUFA含量会与ω-6PUFA形成吸收竞争,进一步降低ω-6PUFA含量。不同脂肪酸只有在适宜的ω-6/ω-3PUFA下才能发挥有益机体健康的作用[8]。目前,各国及各组织对该比值的推荐值不尽相同,加拿大推荐值为6.25∶1,联合国粮农组织(FAO)与世界卫生组织(WHO)的推荐值为(5~10)∶1,中国营养学会推荐值为(4~6)∶1。当前,我国人民食用植物油主要有大豆油、花生油、菜籽油和玉米胚芽油,其脂肪酸组成中属于ω-6PUFA的亚油酸含量较高,可达脂肪酸总量的一半以上,从而导致ω-6/ω-3PUFA比值严重超出中国营养学会推荐水平[25]。猪肉作为膳食脂肪酸重要来源,其脂肪酸组成的合理性对人类健康具有重要意义。本试验通过在生长育肥猪日粮中添加膨化亚麻籽,将ω-6/ω-3PUFA比值降至中国营养学会推荐范围。Coates等[26]对比了健康人群分别食用普通猪肉和富ω-3PUFA猪肉12周后血清中脂肪酸的组成发现,食用富ω-3PUFA猪肉可以提高正常人体红细胞ω-3PUFA含量,降低血清甘油三酯和血栓素水平,从而达到降低心血管风险的作用。
3.4不同饲喂程序对生长育肥猪血清生化指标和免疫指标的影响
动物血清中TP和ALB含量主要受日粮类型和营养水平影响,可以一定程度上反映机体蛋白质吸收、合成与分解状况。本试验结果显示,各组间血清TP和ALB含量无显著差异,说明屠宰前分别饲喂生长育肥猪膨化亚麻籽日粮30、60、90d对其蛋白质代谢能力均无影响。动物血清中ALT、AST和ALP活性反映了其肝脏功能的受损程度和抗应激能力。本试验中各组间血清ALT、AST和ALP活性无显著差异,说明屠宰前分别饲喂生长育肥猪膨化亚麻籽日粮30、60、90d不会造成应激而影响机体健康。动物机体中血清免疫球蛋白和补体的含量与其免疫能力呈正相关,免疫球蛋白主要分为IgG、IgM、IgA、IgD和IgE5种[27],激活补体途径中含量最多、最重要的成分为C3和C4。陈静等[18]研究发现,日粮中添加1.5%亚麻籽油饲喂杜×长×大38d后可以显著提高其血清IgM水平。本试验结果也显示,屠宰前饲喂亚麻籽日粮60d可以显著提高其血清中C3和C4以及IgM和IgG含量。饲喂膨化亚麻籽日粮的生长育肥猪体内同时存在高水平ω-3PUFA与低ω-6/ω-3PUFA比值引起的免疫功能增强作用[28]和PUFA产生的过氧化毒性作用,各组间免疫功能的差异可能是上述两种作用的不同表现。这表明屠宰前饲喂亚麻籽日粮90d不会影响生长育肥猪的机体健康,且适宜的屠宰前饲喂时间反而有增强免疫功能的效果。
4结论
在本试验条件下,在屠宰前90d开始饲喂添加5%膨化亚麻籽的日粮可以在不影响生长肥育猪生长性能、屠宰性能性能和机体免疫功能,并可显著提高背最长肌和背膘ω-3PUFA含量,使ω-6/ω-3PUFA比值达到中国营养学会推荐范围。
参考文献
[1]Delion S, Chalon S, Guilloteau D, et al. Alpha-linolenic acid dietary deficiency alters age-related changes of dopaminergic and serotoninergic neurotransmission in the rat frontal cortex[J]. J Neurochem, 2010, 66(4): 1582-1591.
[2]Riediger N D, Othman R A, Suh M, et al. A systemic review of the roles of n-3 fatty acids in health and disease[J]. J Am Diet Assoc, 2009, 109(4): 668-679.
[3]Mozaffarian D, Wu J. Omega-3 fatty acids and cardiovascular disease: effects on risk factors, molecular pathways, and clinical events[J]. J Am Coll Cardiol, 2011, 58(20): 2047-2067.
[4]Von Schacky C. Omega-3 fatty acids in cardiovascular disease-an uphill battle[J]. Prostag Leukotr Ess, 2015, 92: 41-47.
[5]Simopoulos A P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases[J]. Exp Biol Med, 2008, 233(6): 674-688.
[6]Ponnampalam E N, Sinclair A J, Holman B W B. The sources, synthesis and biological actions of omega-3 and omega-6 fatty acids in red meat: an overview[J]. Foods, 2021,10(6): 1358.
[7]Ma X, Jiang Z, Lai C. Significance of increasing n-3 PUFA content in pork on human health[J]. Crit Rev Food Sci, 2016, 56(5): 858-870.
[8]Artemis S. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity[J]. Nutrients, 2016, 8(3): 1-17.
[9]Czernichow S, Thomas D, Bruckert E. n-6 Fatty acids and cardiovascular health: a review of the evidence for dietary intake recommendations[J]. Brit J Nutr, 2010, 104(6): 788-796.
[10]Shetty S S, Kumari N S, SHETTY P K. ω-6/ω-3 fatty acid ratio as an essential predictive biomarker in the management of type 2 diabetes mellitus[J]. Nutrition, 2020, 79-80: 110968.
[11]Huang C, Chiba L I, Bergen W G. Bioavailability and metabolism of omega-3 polyunsaturated fatty acids in pigs and omega-3 polyunsaturated fatty acid-enriched pork: a review[J]. Livest Sci, 2021, 243: 104370.
[12]Corino C, Rossi R, Cannata S, et al. Effect of dietary linseed on the nutritional value and quality of pork and pork products: systematic review and meta-analysis[J]. Meat Sci, 2014, 98(4): 679-688.
[13]邓波, 门小明, 吴杰, 等. 亚麻籽对生长育肥猪生长性能、胴体性状、肉质和脂肪酸组成的影响[J]. 动物营养学报, 2019, 31(9): 4024-4032.
[14]吴永保, 杨凌云, 闫海洁, 等. 饲粮中添加微藻和亚麻籽提高鸡蛋黄中ω-3 多不饱和脂肪酸含量对比研究[J]. 动物营养学报, 2021, 27(10): 3188-3197.
[15]Burnett D D, Legako J F, Phelps K J, et al. Biology, strategies, and fresh meat consequences of manipulating the fatty acid composition of meat[J]. J Anim Sci, 2020, 98(2): 1-12.
[16]Vehovsky K, Zadinova K, Stupka R, et al. Fatty acid composition in pork fat: de-novo synthesis, fatty acid sources and influencing factors-a review[J]. Agron Res, 2018, 16(5): 2211-2228.
[17]Nguyen D V, Malau-Aduli B S, CAVALIERI J, et al. Supplementation with plant-derived oils rich in omega-3 polyunsaturated fatty acids for lamb production[J]. Vet Anim Sci, 2018, 6: 29-40.
[18]陈静, 刘显军, 王彤, 等. 饲粮必需脂肪酸组成对育肥猪生长性能和血清生化指标的影响[J]. 动物营养学报, 2019, 31(8): 3543-3550.
[19]石宝明, 郎婧, 单安山. 亚麻籽和维生素 E 对育肥猪胴体脂肪酸构成的影响[J]. 中国畜牧杂志, 2012, 48(13): 53-56.
[20]Kouba M, Enser M, Whittington F M, et al. Effect of a high-linolenic acid diet on lipogenic enzyme activities, fatty acid composition, and meat quality in the growing pig[J]. J Anim Sci, 2003, 81(8): 1967-1979.
[21]Hallenstvedt E, Kjos N P, Verland M. Changes in texture, colour and fatty acid composition of male and female pig shoulder fat due to different dietary fat sources[J]. Meat Sci, 2012, 90(3): 519-527.
[22]Hallenstvedt E, Kjos N P, Rehnberg A C, et al. Fish oil in feeds for entire male and female pigs: changes in muscle fatty acid composition and stability of sensory quality[J]. Meat Sci, 2010, 85(1): 182-190.
[23]Nuernberg K, Fischer K, Nuernberg G, et al. Effects of dietary olive and linseed oil on lipid composition, meat quality, sensory characteristics and muscle structure in pigs[J]. Meat Sci, 2005, 70(1): 63-74.
[24]李德发.中国猪营养需要[M]. 北京: 中国农业出版社, 2019.
[25]魏永生, 郑敏燕, 耿薇, 等. 常用动、植物食用油中脂肪酸组成的分析[J]. 食品科学, 2012, 33(16): 188-193.
[26]Coates A M, Sioutis S, Buckley J D, et al. Regular consumption of n-3 fatty acid-enriched pork modifies cardiovascular risk factors[J]. Brit J Nutr, 2009, 101(04): 592-597.
[27]程远之, 周洪彬, 虞财华, 等. 不同类型酸化剂对断奶仔猪生长性能和免疫功能的影响[J]. 动物营养学报, 2021, 33(5): 2575-2584.
[28]Simopoulos A P. The importance of the ratio of omega-6/omega-3 essential fatty acids[J].Biomed. Pharmacother, 2002, 56:365-379.
文章摘自:许明曙,华颖,刘国良,林豪,于震,陈玉,李俊跃,王腾浩.不同膨化亚麻籽日粮饲喂程序对生长育肥猪生长性能、脂肪酸沉积和免疫功能的影响[J/OL].中国畜牧杂志:1-12[2022-01-12].
