摘 要:为探究外源硒肥对胡麻产量、农艺性状、生理特性及品质的调控作用,本研究以‘同亚16’(TY16)和‘晋亚10号’(JY10)两个胡麻品种为试验材料,通过田间试验设置六个不同硒肥剂量:清水对照(T0)、20 g/hm2 NaSeO3(T1)、30 g/hm2 NaSeO3(T2)、50 g/hm2 NaSeO3(T3)、70 g/hm2 NaSeO3(T4)、90 g/hm2 NaSeO3(T5)。硒肥在胡麻盛花期和青果期叶面喷施硒肥两次。研究分析了硒肥对胡麻产量及产量构成因子、光合作用参数、抗氧化酶活性、丙二醛含量、籽粒硒含量和品质的影响。结果表明,50 g/hm2的叶面喷施显著提升了TY16和JY10的产量,较对照组分别增加了9.91%和12.51%,主要得益于单株果数和每果粒数的增加。叶片的净光合速率、气孔导度、蒸腾速率等光合参数以及叶绿素含量相比对照组有显著提升,表明光合作用得到增强。超氧化物歧化酶(SOD)活性增幅为4.40%~20.12%,同时显著降低了丙二醛含量,减轻了叶片膜脂过氧化损伤。在T5处理组中,TY16和JY10籽粒硒含量达到最高,分别为0.461 mg/kg、0.505 mg/kg,比对照组提高18.47和21.96倍。在品质方面,施用硒肥后籽粒中的亚油酸含量显著增加5.49%~24.56%,TY16和JY10的亚麻酸含量较对照组(T0)分别上升6.96%和5.84%。因此,50 g/hm2硒肥施用不仅提高了胡麻的产量和农艺性状,还增强了光合作用和叶绿素含量,提升了植株抗氧化能力,并通过增加籽粒硒含量和不饱和脂肪酸亚油酸与亚麻酸的含量,显著提高了胡麻籽的营养价值。
关键词:胡麻;叶面喷硒;光合作用;产量;品质
胡麻(Linum usitatissimum L.)作为我国五大油料作物之一[1],截至2020年,全国种植面积约19.3万hm2,年产量约28.4万t,在保障国家的食用油安全和工业用油需求中发挥了重要作用。胡麻油以其独特的营养成分越来越受到人们的广泛关注,其富含α-亚麻酸等多种不饱和脂肪酸在人体内可转化为对健康至关重要的DHA(Docosahexaenoic acid,二十二碳六烯酸)和EPA(Eicosapentaenoic acid,二十碳五烯酸)[2],这些Omega-3脂肪酸对于促进大脑发育、增强认知功能、维护心血管健康以及预防相关疾病具有极其重要的作用[3,4]。胡麻还富含有其他重要营养活性成分,如亚麻子多糖、亚麻木酚素、蛋白质、膳食纤维等。经常食用胡麻子,具有降血脂、降血压、益智、保护视力、提高人体免疫力等功效[5,6]。因此,胡麻不仅可以用于胡麻油的制取,还可以胡麻子粉等形式添加到食品中,从而起到提高食品营养价值和改善食品风味的作用。
硒(Se)是人体必需的微量元素,具有多种生物学功能,被誉为“抗癌之王”和“长寿元素”,其丰缺直接关系到人体健康,在心血管疾病、甲状腺疾病、糖尿病、儿童自闭症等防治方面具有积极的辅助作用[7,8,9]。同时硒在植物体内也是不可缺少的微量有益元素,在提高植物光合作用及抗氧化能力、促进作物生长等方面具有重要作用,一定程度上还能增加作物产量[10,11]。我国是世界上缺硒最严重的地区,全国低硒地区占总面积的72%[12],人均日硒摄入量为26~32μg/d,远不足世界卫生组织推荐值(50~200μg/d)的下限[13]。饮食摄入硒是人体补充硒的主要途径,粮食作物作为饮食硒的重要来源之一,然而大多数农产品硒含量均不足60μg/kg[14,15],无法满足人们的日常需求。因此,提高作物中硒浓度的生物强化技术,增加富硒作物的生产尤为必要。
本研究以同亚16号和晋亚10号为材料,在盛花期、青果期两次叶面喷施不同剂量的亚硒酸钠,研究硒肥对胡麻产量及产量因子、株高和茎粗等农艺形状、光合作用指标、抗氧化酶、籽粒含硒量和品质的影响,旨在寻找最合适的胡麻外源硒喷施的最佳浓度,为解决我国居民的硒缺乏问题提供一种农业技术解决方案,为富硒胡麻的栽培提供理论依据和技术支持。
1 材料和方法
1.1 试验材料
在前期预试验基础上,供试品种选用同亚16(山西农业大学提供)和晋亚10号(山西农业大学提供)。试验地前茬为高粱,试验土壤(0~20cm)pH为8.22,有机质为11.3g/kg,碱解氮为39.9mg/kg,有效磷为11.8mg/kg,速效钾为102mg/kg,硒0.11mg/kg。
1.2 试验方法
试验于2023年在山西省吕梁市柳林县三角镇靳家山进行,选取籽粒饱满、无破损的种子于4月15日随耕播种,铁耧条播,行距25cm,播深3~4cm。试验采取随机区组设计,共设置6个硒肥剂量,分别是清水对照(T0),10g/hm2 NaSeO3(T1)、30g/hm2 NaSeO3(T2)、50g/hm2 NaSeO3(T3)、70g/hm2 NaSeO3(T4)、90g/hm2 NaSeO3(T5),重复3次,小区面积6m×6m。分在胡麻盛花期和青果期叶面喷施硒肥两次,采用小型手动喷雾器将硒肥均匀地喷在胡麻植株上。喷施硒肥选择在无风的晴天,喷施时间在上午10:00前,其他管理按大田常规措施管理。在90%的胡麻蒴果变为黄褐色时,人工收获、脱粒。
1.3 测定项目与方法
1.3.1 农艺性状
成熟期在每个小区随机选取30株胡麻测定农艺性状,包括株高、工艺长度、茎粗。株高,自子叶痕迹量至蒴果顶端;工艺长度,子叶迹至主茎第一分枝处的长度;茎粗,主茎种间部位直径,使用游标卡尺测量。
1.3.2 小区测产
胡麻成熟期各小区采样30株进行室内考种,分别测定单株果数、每果粒数、千粒重和单株产量。收获按小区单打单收,晒干后称取胡麻籽粒质量,测得小区实际产量。
1.3.3 光合参数测定
青果期喷施硒肥一周后测定植株光合参数,提前选定各处理组的5株长势一致的植株挂牌标记。在晴天上午9:00至11:00使用CI-340光合测定仪(CID Bio-Science,美国)测定倒二叶中部净光合速率(Pn)、气孔导度、蒸腾速率及胞间CO2浓度,计算平均值。
1.3.4 抗氧化酶和丙二醛测定
各个处理取3个生物学重复,称取1 g叶片样本冰浴研磨,使用检测试剂盒(苏州研犀生物科技有限公司,中国),按照说明书测定超氧化物歧化酶(SOD)活性何和丙二醛(MDA)含量。
1.3.5 硒含量测定
硒含量使用氢化物发生-原子荧光法测定,称取固体试样0.1 g,置于聚四氟乙烯消解内罐中,加硝酸5mL浸泡过夜。盖好内盖,旋紧不锈钢外套,放入恒温干燥箱,80℃保持1~2h,120℃保持1~2h,再升至160℃保持4h,在箱内自然冷却至室温,打开后在电热板上加热赶酸近干,加5ml盐酸溶液(6mol/L)溶解1mL铁氰化钾(100g/L),转移至25ml容量瓶中,用纯水定容,混匀待测。
1.3.6 亚油酸和亚麻酸含量
成熟期收获后,每小区取胡麻籽粒50g,委托苏州研犀生物科技有限公司(苏州,中国)使用气相色谱仪(Thermo Scientific ISQ 7610,美国)测定籽粒亚麻酸和亚油酸含量。
1.4 数据处理
采用Excel 2010进行分析和整理,使用SAS进行方差分析,用HSD检验法进行差异显著性检验(P<0.05)。
2 结果与分析
2.1 外源硒对胡麻农艺性状的影响
由表1可知,叶面施用硒肥对胡麻的株高、工艺长度和茎粗3个农艺性状均产生显著影响,不同品种对硒肥的响应存在差异。随着硒肥喷施浓度的提高,2个品种的株高呈现出先升高后降低的趋势。在TY16品种中,T3处理组的株高最高,较对照组T0、T1、T2、T4、T5分别提高6.80%、2.73%、1.77%、2.94%和6.10%,并与T0、T5存在显著差异(P<0.05)。在晋亚10号品种中,T3和T4株高较对照组显著提高5.13%~5.18%(P<0.05)。这表明适宜的硒肥浓度能够有效促进胡麻株高的增长。对于工艺长度,TY16品种在T3处理组中最长,随着硒肥浓度增加,工艺长度逐渐减少。JY10号品种也显示类似趋势,其中T3工艺长度最长,分别比T0、T1和T5提高14.29%、13.96%和9.24%,均达显著差异水平(P<0.05)。茎粗的数据表明,两品种的茎粗也随着硒肥浓度的增加呈先升高后降低的趋势,均表现为T3处理组的茎粗最大,较其他处理组提高0.52%~10.99%,这表明喷施硒肥有助于胡麻茎粗的增加,但硒肥浓度过高会对茎粗产生抑制作用。
表1 硒肥对胡麻农艺性状的影响
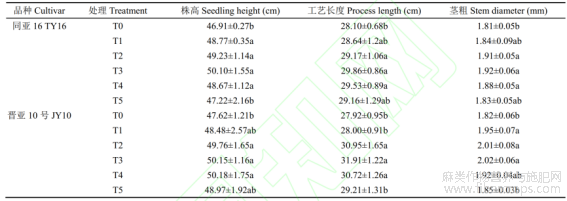
不同小写字母表示处理间差异显著(P<0.05),下同。
2.2 外源硒对胡麻产量的影响
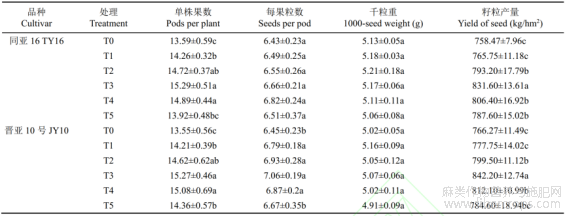
由表2可知,叶面施用硒肥对胡麻的产量及产量构成均有显著影响。随着叶面喷施硒肥浓度的增加,两个胡麻品种产量呈现先升高后降低的趋势,T3浓度的胡麻籽粒产量最高。TY16的T3处理组产量较T0、T1、T2、T4和T5依次提高9.64%、8.60%、4.84%和3.13%,差异均达显著水平(P<0.05);JY10籽粒产量在T3处理组中达到最高(842.2kg/hm2),较T0、T1和T5分别提高9.91%、8.29%和5.59%,呈显著差异(P<0.05)。这表明硒肥对提高胡麻产量效果显著。分析产量构成因素表明,随着硒肥浓度的增加,TY16单株果数在T3处理组达到最大值,分别较T0、T1和T5增加12.51%、7.22%和9.84%,均达统计学差异(P<0.05)。与TY16品种相似,JY10号品种的单株果数也随着硒肥浓度的增加而增加,T3处理组中单株果数最高,较T0、T5显著提高12.69%和6.34%(P<0.05),表明适宜浓度的硒肥有利于提高胡麻的单株果数。每果粒数方面,TY16在T4处理组中达到最高,较其他处理组的增幅为2.40%~6.07%。与TY16不同,JY10每果粒数在T3浓度时最大,较T0、T5分别增加9.46%和5.85%,差异达显著水平(P<0.05)。两品种的千粒重在T1和T2处理组中相对较高,但各处理组间变化均未达显著差异(P>0.05),这可能表明千粒重的遗传因素较强,受硒肥浓度的影响较小。以上结果说明,适宜浓度硒肥可以显著提高胡麻产量,主要归因于单株果数和每果粒数的提高。
表2 硒肥对胡麻产量及产量构成的影响
2.3 外源硒对胡麻光合特性和SPAD的影响
由图1可知,硒肥处理对胡麻光合特性具有显著的促进作用。随着硒肥浓度的增加,两品种T3处理组的净光合速率达到最大值(图1A),TY16品种的T3处理组较T0、T1、T2、T4和T5分别增加44.86%、24.65%、14.04%、21.13%和37.79%,JY10号品种的T3净光合速率较T0、T1、T2、T4和T5依次提高57.69%、38.61%、17.70%、14.94%和25.52%,均达显著差异水平(P<0.05)。气孔导度也呈现类似趋势,两品种均在T3处理组达到最高值(图1B),其中TY16的T3气孔导度较其他处理组的增幅达5.86%~24.29%,JY10号的T3较其他处理组增幅为7.22%~30.84%,均与T0、T1和T2间差异性显著(P<0.05)。两品种的蒸腾速率在T3浓度也达到最大值(图1C),表明适宜硒肥浓度有利于增强胡麻水分调节能力从而促进植株光合作用能力。而在T4和T5处理组蒸腾速率显著下降,这可能是过高硒肥浓度对植株产生不利影响。与以上光合特征参数不同,叶片胞间CO2浓度随着硒肥浓度的增加呈先降低后逐渐上升趋势(图1D)。两品种T3处理组的胞间CO2浓度最低,较其他处理组降幅达7.66%~24.54%,与T0、T1和T5差异性均达显著水平(P<0.05)。JY10号品种在T3处理组中净光合速率高于TY16,而胞间CO2浓度更低,这表明JY10可能更高效地利用硒肥来提高其光合效率。
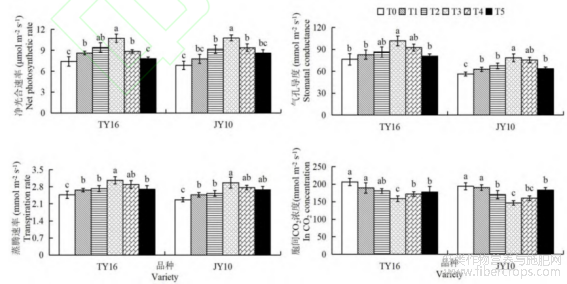
图1 硒肥对胡麻光合作用的影响
不同小写字母表示处理间差异显著(P < 0.05),下同。
由图2可知,硒肥处理能够显著提高胡麻的SPAD值。在两个胡麻品种中,随着硒肥浓度的增加,SPAD值呈现上升趋势,于T3处理组达到最大值。TY16的T3处理组SPAD值较T0、T1、T2和T5分别提高10.85%、7.54%、5.46%和5.62%,差异均达显著水平(P<0.05)。JY10号品种T3处理组的SPAD较其他处理组增幅为3.24%~12.32%。两品种SPAD值在T4和T5均有所下降,但仍高于对照组。对比品种在相同硒肥下的表现,JY10号品种SPAD值均不同程度高于TY16品种,这可能是JY10号对硒肥有更好的利用效率,能够更好地促进光合作用和叶绿素合成。
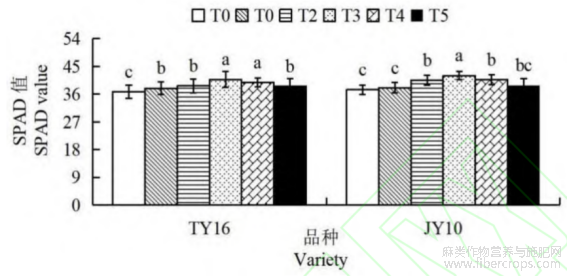
图2 硒肥对胡麻SPAD值的影响
2.4 外源硒对胡麻抗氧化酶和丙二醛的影响
由图3可知,硒肥处理对胡麻的抗氧化酶活性和丙二醛含量有显著影响。在两个品种中,随着硒肥浓度的增加,抗氧化酶活性呈先上升后迅速降低趋势。两品种的SOD活性均在T3处理组达到最高值(图3A),TY16的T3处理组SOD活性较T0、T4和T5分别提高10.84%、8.97%和20.12%,均表现为显著差异(P<0.05);JY10号T3处理组的SOD活性较其余处理组增幅为4.40%~14.15%,显著高于T0、T1、T4和T5(P<0.05)。丙二醛含量变化趋势则与抗氧化酶活性趋势相反。两品种的丙二醛含量随硒肥浓度增大逐渐降低,均在T3处理组达到最低值(图3B)。TY16的T3处理组丙二醛含量较T0、T1和T5显著降低11.36%、10.34%和15.22%(P<0.05);JY10号的T3处理组较其余处理组降幅为4.82%~10.23%,均达显著差异水平(P<0.05)。以上结果说明,适当的硒肥浓度可以提升胡麻抗氧化酶活性,有助于减少氧化损伤。
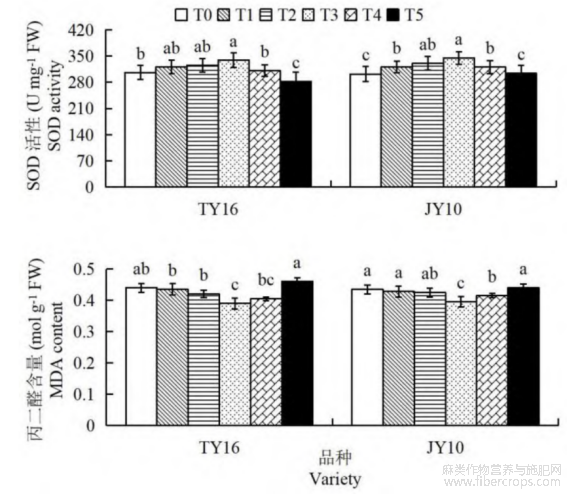
图3 硒肥对胡麻SOD活性和丙二醛含量的影响
2.5 外源硒对胡麻籽粒硒含量的影响
由图4可知,施用外源硒肥显著提高了胡麻籽粒硒含量。随着硒肥浓度的增加,两个品种的籽粒硒含量均呈上升趋势,而增速逐渐降低。TY16和JY10号胡麻籽粒硒含量在T5处理组中达到最高值,依次为0.461mg/kg、0.505mg/kg,分别是各自T0对照组籽粒硒含量的18.47和21.96倍,均达到显著差异水平(P<0.05)。在相同硒肥处理下,JY10号品种的籽粒硒含量普遍高于TY16品种,这可能是JY10号品种对硒肥有更好的吸收及积累能力。
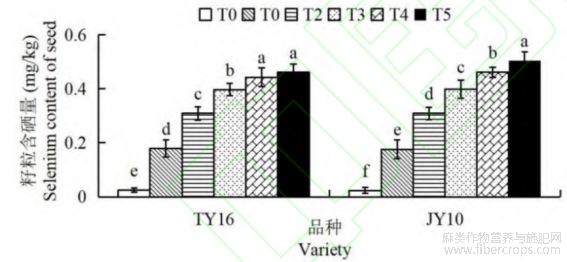
图4 硒肥对胡麻籽粒硒含量的影响
2.6 外源硒对胡麻亚油酸和亚麻酸含量的影响
由图5可知,叶片喷施硒肥对胡麻籽粒的亚油酸和亚麻酸含量有显著影响。随着硒肥浓度的增加,两品种的亚油酸和亚麻酸含量呈现出上升后趋于稳定趋势。TY16品种的T4处理组亚油酸含量最高,分别较T0、T1组提高24.53%、20.31%,呈显著差异水平(P<0.05)。JY10号T4处理组的亚油酸含量比T0、T1、T2和T5 依次增加25.63%、18.71%、11.36%和5.49%,均表现统计学差异(P<0.05)。两品种的亚麻酸含量均在T4处理组达到最高,TY16品种T4较其他处理组的增幅为1.26%~5.84%,与T1、T2间均表现为显著差异(P<0.05)。JY10号亚麻酸含量在T4处理中也达到最高,较T0、T1和T5显著提高6.96%、5.83%和4.70%(P<0.05)。在相同硒肥处理下,JY10号品种的亚油酸和亚麻酸含量普遍高于TY16品种,表明JY10号能够更有效地利用硒肥来提高油脂品质。
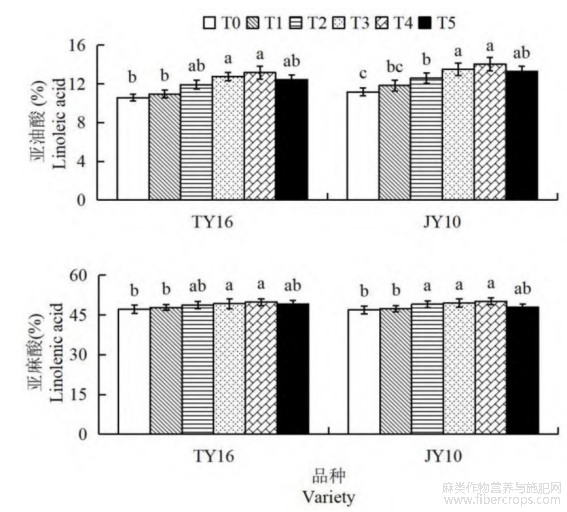
图5 硒肥对胡麻亚油酸和亚麻酸含量的影响
3 讨论
硒作为一种对人体有着重要作用的微量元素,对作物的产量同样产生显著影响。姜宗昊等[16]研究发现,土壤施加高硒矿粉显著提高了小麦的千粒重和穗粒数,其中周黑麦1号品种产量增幅26.42%。石吕等[17]研究表明,水稻灌浆期喷施100 g/hm2浓度的硒肥对千粒重和结实率有明显促进作用,成熟期干物质积累量增加。岳琳祺等[18]研究发现,谷子抽穗期施用110 g/hm2硒肥显著提高了穗粒重和穗重等产量构成因子。本研究表明:施用50 g/hm2硒肥后胡麻产量较对照增幅达9.64%~9.91%,主要归因于单株果数和每果粒数显著提高。这可能是硒肥通过影响植株激素的平衡,促进了生长素(IAA)和赤霉素(GA)的合成与调节,直接影响花器官的发育和果实的形成,从而增加了单株果数和每果粒数[19]。此外,施用硒还可能影响植株对土壤氮、磷和钾等主要营养元素的吸收和转运,这些都是影响胡麻产量的重要营养素。施用适宜浓度的硒肥还能够显著促进植物的生长发育,并对其形态特征产生积极的影响[20]。彭晓伟等[11]研究表明,随着施硒量的增加,冀谷19号和晋谷19号株高、茎粗均不同程度增加。本研究表明,50 g/hm2硒肥显著提高了胡麻株高和茎粗等农艺性状。这可能是硒通过调节植物体内的激素平衡并优化植株的形态建成,影响细胞的伸长和分裂,促进植株的生长发育。同时,本研究表明,过量施用硒肥会对胡麻产生毒害作用,反而抑制植株生长,与余江敏等[21]研究结果一致。因此,在农业生产中需要精确控制硒肥的施用量及方式,对不同土壤和气候条件下的硒肥施用进行精细调整,以实现胡麻产量和效益的最大化。
光合作用作为能量转换和生长发育的核心过程,硒元素已在多项研究中被证实能够显著影响植物的光合作用效率[22]。Wang等[23]研究发现,在低硒浓度(2.5 mg/kg)下,玉米净光合速率提高了47.47%,蒸腾速率(Tr)增加了63.46%。Ulhassan等[24]研究表明,甘蓝叶面喷施氨基酸螯合硒处理,有效改善气体交换参数和降低胞间CO2浓度来增强光合作用。本研究结果表明,T3处理组硒肥浓度显著促进了胡麻的光合作用,净光合速率显著提高44.86%,与Xu等研究结果一致。这可能是硒肥通过增强光合电子传递链中光系统II(PSII)的稳定性,提高光能转换效率,从而提高光合作用。Das等[25]在谷类作物中发现,硒显著提升了幼苗气孔导度和蒸腾速率,这与本研究中气孔导度和蒸腾速率在T3组达到最大值的结果一致。这可能是硒肥改善了气孔开闭调控机制,优化胡麻的气体交换能力及水分利用效率,进一步证实硒在调节植物水分关系和气体交换过程中的作用[26]。本研究表明,T3处理组的胞间CO2浓度显著低于其他处理组,降幅达24.54%。这一现象可能是硒肥促进了关键光合酶Rubisco的表达或活性,保护该酶免受氧化损伤,加速了Calvin循环中催化CO2固定成有机物,进而降低了胞间CO2浓度。叶绿素作为光合作用中的关键色素,其含量的变化会直接影响植物的光合作用效率和生长发育状态[27]。薛竟一等[28]研究表明,叶面喷施1 mg/L硒处理下的葡萄叶绿素含量最高,比对照组提高24.20%。本研究表明,配施50 g/hm2 NaSeO3显著提升了胡麻叶绿素含量,较对照组增加12.32%,这与余江敏等[29]研究结果一致。这可能是硒通过调节叶绿素生物合成途径中的关键酶活性来增加叶绿素含量。特别是,硒与巯基相关的抗氧化酶(如:谷胱甘肽过氧化物酶)的活性可能在此过程中发挥作用。这种酶类通常参与抗氧化过程,有助于维护细胞的还原状态,从而直接或间接地促进叶绿素的生物合成,最终提高叶绿素含量。
抗氧化酶系统是植物细胞内抵御活性氧(ROS)诱导的氧化损伤的主要防线,硒元素对提高植物的抗氧化能力具有积极作用[30]。张岩松等[31]研究表明,喷施适宜浓度硒肥(1.0 mg/L)后,生菜叶片的抗氧化酶活性均不同程度提高,SOD活性较对照组增幅达1.54倍,丙二醛含量显著下降36%。Wang等[32]研究发现,经硒肥处理后鲜切苹果表现出较高的SOD和CAT 活性,膜脂过氧化程度显著降低,有效缓解了鲜切苹果在储存阶段的褐变现象。本研究表明,随着硒肥浓度增加,叶片中SOD活性首先升高并随后下降,在T3处理组SOD活性达到峰值,丙二醛(MDA)含量也显著降低。SOD作为植物体内一种关键的抗氧化酶,负责将超氧自由基(O2-)催化歧化为氧分子(O2)和过氧化氢(H2O2)的功能。硒肥通过提高SOD及其他抗氧化酶的活性,增强了植物细胞清除过量ROS的能力,这一机制对于维护细胞内氧化还原平衡具有至关重要的作用[33]。硒还通过增强细胞的抗氧化能力,显著降低了氧化应激对细胞结构与功能的影响,特别是在保护生物膜方面表现显著,通过抑制丙二醛MDA的产生,有效减轻了膜脂过氧化的损伤[34]。但喷施硒肥浓度超过特定的临界阈值时,其累积的毒性作用可能导致植物体内抗氧化酶系统的活性受抑,削弱其清除ROS的能力,导致细胞膜脂质过氧化程度加剧。因此,在农业生产中应用硒肥时必须严格控制其浓度范围,以避免引起硒的潜在毒性效应,确保其对作物生长和抗氧化系统的积极影响。
富硒产品通过补充人体所需的微量元素硒,不仅能够显著增强免疫力、预防由硒缺乏引发的相关疾病,还参与调节体内的氧化应激反应,有效延缓细胞衰老过程[35]。De等[36]研究表明,随着土壤中施硒量增加,小麦籽粒、秸秆和根系干物质中硒含量显著增加,其中籽粒硒含量由0.155 mg/kg显著提升至0.732 mg/kg。本研究表明,随着施用硒肥浓度增加,胡麻籽粒中硒含量呈上升趋势,籽粒硒含量最高为0.505 mg/kg,达到富硒农产品标准中油料作物类硒含量要求。本研究还发现,当硒肥浓度超过70 g/hm2时,籽粒硒含量增加幅度显著降低,这与冯学金等[37]研究结果基本一致。胡麻籽粒作为多种人体必需脂肪酸的宝贵来源,尤其富含的亚油酸和亚麻酸无法通过人体自身的代谢途径合成,因此必须通过饮食摄入来满足生理需求[38]。本研究中,硒肥对胡麻籽中亚麻酸和亚油酸含量的显著提高,其中亚油酸较对照组显著提高24.53%。类似的生理效应也在Li等[39]研究中观察到,其中纳米硒被发现能够增强辣椒和芹菜中的相应生理成分。特别是在辣椒中,纳米硒通过促进植物体内苯丙烷途径与脂肪酸途径的相互作用,显著提高了脂肪酸代谢通路中关键基因和化合物的表达量。这些发现表明,硒肥可能通过调节植物的代谢过程,对胡麻籽的脂肪酸组成产生积极影响。此外,硒还作为一种强效的抗氧化剂,对增强植物的抗氧化防御系统、保护脂肪酸合成相关酶免受氧化损伤起着重要作用,从而有利于亚麻酸和亚油酸的生物合成[40]。因此,硒的作用不仅局限于其直接的生化活性,还可能通过影响植物激素调节和代谢通路的整体动态来增强脂肪酸的合成。综上,硒肥的施用显著提升了胡麻籽的营养价值和功能特性,表明硒肥的应用在农业生产中具有重要的实际意义和潜在的经济价值。
4 结论
胡麻盛花期和青果期叶面喷施50g/hm2硒肥可显著增加胡麻产量,主要归因于单株果数和每果粒数的提高。农艺形状方面,硒肥可以对胡麻的株高和茎粗有明显促进作用,但促进效果随硒肥浓度的提高而迅速降低。在光合作用方面,硒肥通过提高叶片叶绿素含量、净光合速率、气孔导度和蒸腾速率,以及显著降低胞间二氧化碳浓度,增强了胡麻的光合能力及有机物质的积累。这些变化可能与硒肥提高抗氧化系统中SOD活性和降低丙二醛含量的作用有关,从而增强了胡麻的抗氧化能力,保护了光合机构免受氧化损伤。硒肥还促进了胡麻籽粒中硒含量的上升,为通过饮食补硒提供了新的途径。在胡麻籽的品质方面,硒肥显著促进了亚麻酸和亚油酸这两种必需脂肪酸的积累,进一步增强了胡麻籽的营养价值和功能特性。接下来将进一步探索硒肥施用的最佳时期及剂量效应关系以及在不同栽培条件下的应用效果,以实现硒肥在可持续农业中的高效利用。
参考文献
[1] Zheng J C,Bin Y,Yu H G,et al. Agronomic cultivation measures on productivity of oilseed flax:A review. Oil Crop Science,2022,7(1):53-62.
[2] 禹晓,邓乾春,黄庆德,等. 亚麻油的制油工艺及其开发利用研究进展. 食品研究与开发,2011,32(1):147-153.
[3] 陈福妮,王卫飞,穆利霞,等. 6种富含α-亚麻酸食用油脂的主要组成成分及消化特征研究进展. 中国油脂,2024,49(1):60-66.
[4] 党照,牛俊义,党占海,等. 胡麻种子发育过程中α-亚麻酸的积累规律. 西北农业学报,2014,23(12):90-95.
[5] Makni M,Fetoui H,Gargouri N K,et al. Antidiabetic effect of flax and pumpkin seed mixture powder: effect on hyperlipidemia and antioxidant status in alloxan diabetic rats. Journal of Diabetes and Its Complications,2011,25(5):339-345.
[6] 邵文捷. 亚麻籽-二十一世纪新型功能性食品. 食品工业,2012,33(12):145-147.
[7] Campos J R,Severino P,Ferreira C S,et al. Linseed essential oil - source of lipids as active ingredients for pharmaceuticals and nutraceuticals. Current Medicinal Chemistry,2019,26(24):4537-4558.
[8] 孙敏,王宇,郭玉成,等. 微量元素硒与儿童孤独症谱系障碍关联的Meta分析. 神经疾病与精神卫生,2022,22(3):158-165.
[9] Liu Q,Zhao X,Ma J,et al. Selenium (Se) plays a key role in the biological effects of some viruses: Implications for COVID-19. Environmental Research,2021,196:110984.
[10] 马俊桃,周文,李静浩,等. 外源硒调控植物重金属胁迫机制的研究进展. 中国农业科技导报,2022,24(6):27-35.
[11] 彭晓伟,蒋沛含,张爱军,等. 叶面喷施硒肥对谷子农艺性状及品质的影响. 山西农业大学学报(自然科学版),2022,42(1):1-9.
[12] Dinh Q T,Cui Z,Huang J,et al. Selenium distribution in the Chinese environment and its relationship with human health: A review. Environment International,2018,112:294-309.
[13] Guerrero B,Llugany M,Palacios O,et al. Dual effects of different selenium species on wheat. Plant Physiology and Biochemistry,2014,83:300-307.
[14] Williams P N,Lombi E,Sun G X,et al. Selenium characterization in the global rice supply chain. Environmental Science & Technology,2009,43(15):6024-6030.
[15] 余琼,张翔,司贤宗,等. 硒在农作物方面的研究进展. 山西农业科学,2018,46(12):2122-2126.
[16] 姜宗昊,刘玉秀,张正茂,等. 土壤不同硒含量对小麦主要产量相关性状和硒吸收利用的影响. 西北农业学报,2021,30(1):41-49.
[17] 石吕,薛亚光,石晓旭,等. 喷施硒肥对富硒土壤水稻产量、品质及硒分配的影响. 中国土壤与肥料,2022,(10):174-183.
[18] 岳琳祺,郭佳晖,白雄辉,等. 叶面喷施硒肥对不同基因型谷子农艺性状及籽粒硒含量的影响. 中国农业科技导报,2021,23(4):154-163.
[19] Rakoczy-Lelek R,Smolen S,Grzanka M,et al. Effectiveness of foliar biofortification of carrot with iodine and selenium in a field condition. Frontiers in Plant Science,2021,12:656283.
[20] Tsivileva O M,Perfileva A I. Mushroom-derived novel selenium nanocomposites' effects on potato plant growth and tuber germination. Molecules,2022,27(14):4438
[21] 余江敏,黄梅燕,杨苛,等. 不同质量浓度外源硒对鲜食玉米硒富集和产量、品质的影响. 作物研究,2022,36(3):207-210.
[22] Luo H,Xing P,Liu J,et al. Selenium improved antioxidant response and photosynthesis in fragrant rice (Oryza sativa L.) seedlings during drought stress. Physiology and Molecular Biology of Plants,2021,27(12):2849-2858.
[23] Wang M,Mu C,Li Y,et al. Foliar application of selenium nanoparticles alleviates cadmium toxicity in maize (Zea mays L.) seedlings: Evidence on antioxidant, gene expression, and metabolomics analysis. Science of the Total Environment,2023,899:165521.
[24] Ulhassan Z,Gill R A,Huang H,et al. Selenium mitigates the chromium toxicity in Brassicca napus L. by ameliorating nutrients uptake, amino acids metabolism and antioxidant defense system. Plant Physiology and Biochemistry,2019,145:142-152.
[25] Das S,Biswas A K. Comparative study of silicon and selenium to modulate chloroplast pigments levels, Hill activity, photosynthetic parameters and carbohydrate metabolism under arsenic stress in rice seedlings. Environmental Science and Pollution Research,2022,29(13):19508-19529.
[26] Jawad H M,Ali R M,Khan I,et al. Selenium and salt interactions in black gram (Vigna mungo L): ion uptake, antioxidant defense system, and photochemistry efficiency. Plants (Basel),2020,9(4):467.
[27] 朱波,徐绮雯,马淑敏,等. 干旱缺钾对油菜内源激素、光合作用和叶绿素荧光特性的影响. 中国油料作物学报,2022,44(3):570-580.
[28] 薛竟一,王瑞雪,魏蓉,等. 叶面喷施亚硒酸钠对葡萄果实品质及叶片衰老的影响. 中国土壤与肥料,2018(6):161-165.
[29] 余江敏,马涛,黄梅燕,等. 叶面喷硒对鲜食玉米籽粒硒富集、叶绿素和糖含量的影响. 陕西农业科学,2023,69(6):9-13.
[30] Das S,Majumder B,Biswas A K. Selenium alleviates arsenic induced stress by modulating growth, oxidative stress, antioxidant defense and thiol metabolism in rice seedlings. International Journal of Phytoremediation,2022,24(7):763-777.
[31] 张岩松,黄皓婷,韩莹琰,等. 硒浓度对高温胁迫下生菜抗氧化酶活性的影响. 北京农学院学报,2021,36(4):39-43.
[32] Wang X,Chang F,Dong Q,et al. Selenium application during fruit development can effectively inhibit browning of fresh-cut apples by enhancing antioxidant capacity and suppressing polyphenol oxidase activity. Journal of Plant Physiology,2023,287:154050.
[33] Araujo M A,Melo A,Silva V M,et al. Selenium enhances ROS scavenging systems and sugar metabolism increasing growth of sugarcane plants. Plant Physiology and Biochemistry,2023,201:107798.
[34] Hawrylak-Nowak B,Matraszek R,Szymanska M. Selenium modifies the effect of short-term chilling stress on cucumber plants. Biological Trace Element Research,2010,138(1/3):307-315.
[35] Malagoli M,Schiavon M,Dallacqua S,et al. Effects of selenium biofortification on crop nutritional quality. Frontiers in Plant Science,2015,6:280.
[36] De V P,Platani C,Fragasso M,et al. Selenium-enriched durum wheat improves the nutritional profile of pasta without altering its organoleptic properties. Food Chemistry,2017,214:374-382.
[37] 冯学金,郭秀娟,杨建春,等. 喷施硒肥对胡麻子粒硒含量、产量及品质的影响. 作物杂志,2019(3):155-157.
[38] 孙雪莲,张波,魏长庆,等. 冷榨胡麻油在加速氧化过程中品质变化的研究. 中国粮油学报,2022,37(7):115-121.
[39] Li D,Zhou C,Zhang J,et al. Nanoselenium foliar applications enhance the nutrient quality of pepper by activating the capsaicinoid synthetic pathway. Journal of Agricultural and Food Chemistry,2020,68(37):9888-9895.
[40] Ahmad I,Younas Z,Mashwani Z U,et al. Phytomediated selenium nanoparticles improved physio-morphological, antioxidant, and oil bioactive compounds of sesame under induced biotic stress. ACS Omega,2023,8(3):3354-3366.
文章摘自:王文霞,畅博凯,夏清,等.叶面喷硒对胡麻生理特性、产量及品质的影响[J/OL].作物杂志:1-11[2024-07-05].http://kns.cnki.net/kcms/detail/11.1808.S.20240611.1716.004.html.
