摘 要:为探究绿色环保的诱抗剂对苎麻生长发育以及抗苎麻夜蛾Arctecoerula特性的影响,选取4种常见且经济的诱抗剂水杨酸、几丁寡糖、壳寡糖1500和壳寡糖1000,分别设置8个浓度处理苎麻后喂食苎麻夜蛾,观察其取食叶面积,在田间处理区半人工释放苎麻夜蛾,观察其为害程度,并在收获时测量苎麻株高和原麻产量。结果表明,4种诱抗剂在适宜浓度下均能显著诱导苎麻对苎麻夜蛾的抗性,田间试验和室内试验结果的一致性较好。其中,80×10-6几丁寡糖稀释液处理对苎麻的诱抗效果最高,苎麻夜蛾为害程度指数仅为8.38,其次是120×10-6壳寡糖1000稀释液处理,为害程度指数为16.71,3.0g/L壳寡糖1500处理和1.2mmol/L水杨酸处理对苎麻的诱抗效果最低,为害程度指数分别为27.21和28.07。4种诱抗剂在适宜浓度下都能显著增加苎麻株高和原麻产量,但各诱抗剂处理的增产量无显著差异,原麻产量以40×10-6壳寡糖1000稀释液处理最高,其次为2.0g/L壳寡糖1500处理,最后为120×10-6几丁寡糖稀释液处理和0.2mmol/L水杨酸处理。以提高苎麻对苎麻夜蛾的抗性和原麻产量为评价标准,4种诱抗剂在田间综合增产和防虫功效应用中优先推荐施用80×10-6~120×10-6几丁寡糖稀释液和40×10-6~80×10-6壳寡糖1000稀释液,其次为2.0~3.0g/L壳寡糖1500和0.8~1.2mmol/L水杨酸。
关键词:苎麻夜蛾;诱导抗虫性;诱导因子;防病促生;防效
苎麻Boehmeria nivea是多年生宿根草本作物,也是优良的天然纤维作物,每年可以收获多次,并且其叶粗蛋白含量高(陈继康等,2020),根系发达且可入药,在饲料、医药及土壤修复和水土保持方面也有着很高的应用价值和潜能(揭雨成,2007;Gong et al.,2017;刘冲等,2020)。但在种植过程中病虫害每年能导致苎麻纤维减产20%以上,如苎麻夜蛾Arcte coerula和根腐病等严重发生时甚至绝收(余永廷等,2011;熊常财等,2012;安晓霞,2014)。苎麻夜蛾是苎麻上主要的食叶害虫,属鳞翅目夜蛾科,又被称为摇头虫和红脑壳虫(曾粮斌等,2013)。该虫主要以幼虫取食苎麻叶片为害,使叶片呈网纹状或缺刻状,暴发为害时全田苎麻叶片能在短期内被蚕食一空,仅剩叶柄及叶脉,使苎麻光合作用迅速衰减或者无法进行,导致被害株生长停滞,多生侧枝,加速纤维老化,严重影响苎麻的产量和品质(王玉娥和王绍华,1987;马继武,1989;Zeng et al.,2016)。
目前,苎麻生产中主要采用化学农药控制苎麻夜蛾为害,但农药的大量使用不可避免产生“3R”问题,同时还会损伤天敌。诱导植物抗性是对生态和环境友好的一种防治方法,可以诱导植物体内抗病虫害防御机制的增强表达(Appu&Muthukrishnan,2014;Hussain et al.,2014),并未涉及到基因本质的转变,具有高度的生物安全性(曾粮斌等,2011;孙凯等,2015)。植物保护素俗称植保素,是植物受病原微生物侵害时自身产生的一类起防卫作用的次生代谢物,其产生速度和积累量与植物的抗病性息息相关。植物诱抗剂是一类能诱导植物通过一系列反应产生植保素的物质,可以是无机盐、有机小分子、生物大分子以及病原细胞壁碎片等(邵素琴和李建中,2002;刘晓丽和娄永根,2018),一般分为寡糖类、蛋白类和糖蛋白类3种。植物诱抗剂的使用具有安全有效、抗谱广的特点,在病虫害防控方面具有广阔的应用前景(Li&Zhu,2013;Mostafanezhad et al.,2014;Ayoubi&Soleimani,2015)。目前,植物诱抗剂已经成熟运用于小麦、玉米、番茄和马铃薯等粮食蔬菜生产中(王宏虬等,2013;秦瑞劼,2018;马赫,2019),能够有效提高作物的抗逆性,并且对产量也有不同程度的提高。关于植物诱抗剂对苎麻的诱抗效果研究极少,仅见安晓霞(2014)使用4种植物诱抗剂处理川苎8号,筛选出水杨酸和壳寡糖1500诱导苎麻抗苎麻夜蛾的效果较好。
本研究分别应用4种不同浓度的植物诱抗剂水杨酸、几丁寡糖和壳寡糖(壳寡糖1500和壳寡糖1000)处理NC03苎麻幼苗,采用室内接虫对峙试验摸索各诱抗剂的适宜浓度,通过室外接虫试验验证所选诱抗剂的有效性,并对处理后的苎麻进行生长和产量的跟踪测定,以期为施用植物诱抗剂诱导苎麻对苎麻夜蛾的抗性、减轻苎麻夜蛾为害程度以及提高苎麻抗逆性和产量提供理论和实践依据。
1材料与方法
1.1材料
供试品种:苎麻品种NC03和中苎一号均为中国农业科学院麻类研究所自行选育品种,于中国农业科学院麻类研究所沅江试验站种植,株行距为0.5m×0.5m,密度约2500蔸/667m2,田间管理采用当地常规措施,为当年二茬麻,生长至15~20cm高时供试。
供试虫源:于中国农业科学院麻类研究所沅江试验站苎麻基地内采集1~2龄苎麻夜蛾幼虫,取回后将其放入高14cm、内直径25cm的养虫缸,用新鲜中苎一号苎麻饲喂,选饥饿5h的4龄苎麻夜蛾幼虫用于试验。
试剂:壳寡糖1500,平均分子量1500,纯度98%,青岛博智汇力生物科技有限公司;壳寡糖1000,平均分子量小于1000,纯度98%,大连中科格莱克生物有限公司;几丁寡糖,平均分子量小于1000,纯度98%,中国科学院过程工程研究所糖生化工程国家重点实验室;水杨酸,上海新浦化工厂有限公司。
仪器:Yaxin-1241叶面积仪,北京雅欣理仪科技有限公司;4BM-260型苎麻剥麻机,中国农业科学院麻类研究所自主研发。
1.2方法
1.2.1苎麻苗诱抗剂处理
在田间用4种诱抗剂处理苎麻NC03,为室内试验提供材料后进行田间试验并跟踪测产。将4种诱抗剂按照前期预试验结果,均设置7个浓度梯度,壳寡糖1500浓度分别为0.25、0.5、1.0、2.0、3.0、4.0和5.0 g/L,壳寡糖1000和几丁寡糖浓度同为20×10-6、40×10-6、60×10-6、80×10-6、120×10-6、160×10-6和200×10-6稀释液,水杨酸浓度分别为0.2、0.4、0.8、1.2、1.6、2.0和3.0mmol/L,均以清水处理为对照。选取长势一致、生长至15~20cm高的苎麻,均匀喷施各处理浓度的诱抗剂,以叶片上诱抗剂液滴开始滴落为止,每个诱抗剂处理浓度设4次生物学重复,1个重复为1个小区,每个小区大小为等长宽3m,试验小区随机区组排列,每个小区种植36蔸苎麻。
1.2.2对峙饲养苎麻夜蛾取食面积的测定
为探索4种诱抗剂对苎麻抗苎麻夜蛾的室内诱导效果,取1.2.1中各诱抗剂喷施处理15d后的苎麻第3~5叶,要求大小比较均一,迅速带回实验室,用剪刀剪下多余叶柄,仅留1cm左右叶柄,在叶柄部位用湿润棉球保湿。分别取大小一致的不同浓度诱抗剂处理叶片各1片放入养虫缸中,然后每个养虫缸中放入10头4龄且大小一致的苎麻夜蛾幼虫,待苎麻夜蛾取食12h后利用Yaxin-1241叶面积仪测定取食前后的叶面积,并计算取食面积比例,每个诱抗剂处理浓度设4次生物学重复。
1.2.3苎麻夜蛾田间为害程度的测定
为进一步验证这4种诱抗剂的诱抗效果,在1.2.1田间试验基础上开展接虫试验,通过在田间半人工释放苎麻夜蛾来测定其为害程度。首先在室内饲养6龄苎麻夜蛾幼虫至化蛹,将蛹置于试验区,每4个小区在田字型中间交叉点放苎麻夜蛾雌、雄蛹2对,待自然羽化后仍其自由选择产卵。待田间苎麻夜蛾孵化并生长至6龄时,参照揭雨成(2007)苎麻夜蛾为害程度分级标准,以取食面积占总叶面积的比例(X)对田间各蔸麻受苎麻夜蛾为害程度进行观察记录,并计算为害程度指数。分级标准:0级,X=0;1级,0<X≤20%;3级,20%<X≤40%;5级,40%<X≤60%;7级,60%<X≤80%;9级,80%<X≤100%。为害程度指数=[∑(为害程度级数×该级别叶片数)]/(调查总叶片数×最高级数)。
1.2.4诱抗剂处理后苎麻株高和原麻产量的测定
对1.2.1中分别喷施4种诱抗剂处理的苎麻统一进行常规的肥水管理,在8月收获时,每个小区随机测量10蔸苎麻的株高,计算平均值,并使用4BM-260型苎麻剥麻机剥取原麻,测定原麻产量。
1.3数据分析
用Excel 2019进行数据整理,使用SPSS 25软件对数据进行统计分析,应用Duncan氏新复极差法进行差异显著性检验。
2结果与分析
2.14种诱抗剂于室内诱导苎麻抗苎麻夜蛾的效果
各浓度壳寡糖1500处理苎麻后,苎麻夜蛾取食面积比例均显著低于对照,3.0g/L处理取食面积比例最低,为20.3%,其次是1.0g/L处理和5.0g/L处理,0.25g/L处理取食面积比例最高,2.0g/L处理较1.0g/L处理高。各浓度壳寡糖1000处理苎麻后,苎麻夜蛾取食面积比例均显著低于对照,80×10-6稀释液处理的取食面积比例最低,为15.3%,其次是120×10-6稀释液处理,20×10-6稀释液处理的取食面积比例最高。各浓度几丁寡糖处理苎麻后,除20×10-6稀释液处理的苎麻夜蛾取食面积比例高于对照外,其余浓度处理的取食面积比例均显著低于对照,其中,80×10-6稀释液处理的取食面积比例最低,为9.7%,其次是120×10-6稀释液处理,说明几丁寡糖20×10-6稀释液处理苎麻后会刺激苎麻夜蛾取食。各浓度水杨酸处理苎麻后,除了0.2mmol/L处理的苎麻夜蛾取食面积比例高于对照外,其余处理的取食面积比例均显著低于对照,其中,0.8mmol/L处理的取食面积比例最低,为20.7%,其次是1.2mmol/L处理,说明0.2mmol/L水杨酸处理苎麻后会刺激苎麻夜蛾取食(图1)。
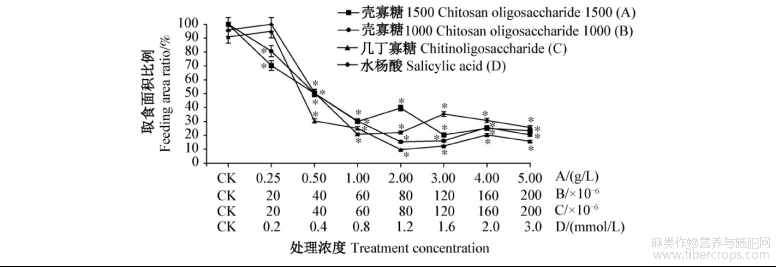
图1 苎麻夜蛾对4种诱抗剂不同浓度处理苎麻叶片的取食面积比例
2.24种诱抗剂于田间诱导苎麻抗苎麻夜蛾的效果
各浓度壳寡糖1500处理苎麻后,苎麻夜蛾田间为害程度指数均显著低于对照,3.0g/L处理的为害程度指数最低,为27.21,其次是1.0g/L处理和5.0g/L处理,0.25g/L处理的为害程度指数最高,2.0g/L处理较1.0g/L处理高,整体趋势与室内试验结果一致。各浓度壳寡糖1000处理苎麻后,苎麻夜蛾田间为害程度指数均显著低于对照,120×10-6稀释液处理的为害程度指数最低,为16.71,其次是80×10-6稀释液处理,与室内试验结果基本一致。各浓度几丁寡糖处理苎麻后,苎麻夜蛾田间为害程度指数均显著低于对照,80×10-6稀释液处理的为害程度指数最低,为8.38,其次是120×10-6稀释液处理;与室内试验结果相比,20×10-6稀释液处理的为害程度指数低于对照,但几丁寡糖最适浓度和诱抗效果并无太大差异。各浓度水杨酸处理苎麻后除0.2mmol/L处理外,苎麻夜蛾田间为害程度指数均显著低于对照,1.2mmol/L处理的为害程度指数最低,为28.07,但0.8~3.0mmol/L浓度处理之间的诱抗效果趋于平稳(图2),整体趋势与室内试验结果一致。
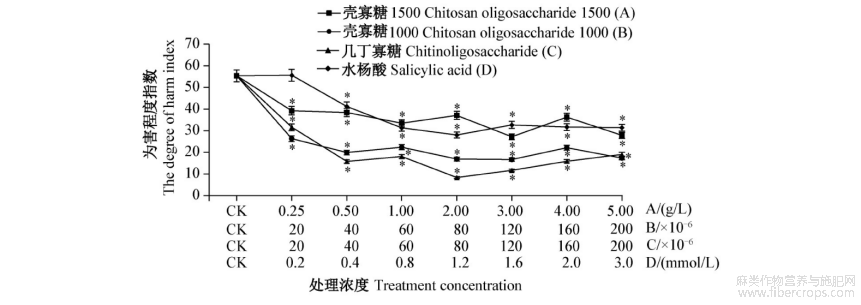
图2 取食4种不同浓度诱抗剂处理苎麻后苎麻夜蛾的田间为害程度指数
2.34种诱抗剂对苎麻株高和原麻产量的影响
4种诱抗剂对苎麻生长和产量均有促进作用,壳寡糖1500和几丁寡糖在较高浓度下增产效果明显,壳寡糖1000和水杨酸在较低浓度下增产效果明显,水杨酸在高浓度时表现出一定的抑制作用,但未达显著水平(图3~4)。
壳寡糖1500处理后,以2.0g/L处理的苎麻株高和原麻产量最高,分别为176.80cm和971.18g,与对照(159.07cm和758.28g)相比分别增加了11.15%和28.18%,3.0g/L处理次之,均显著高于对照,其他浓度处理与对照无显著差异。壳寡糖1000处理后对苎麻株高和原麻产量以40×10-6稀释液处理最高,分别为184.67cm和979.31g,与对照相比分别增加了16.09%和29.15%,20×10-6和60×10-6稀释液处理次之,均显著高于对照,其余浓度处理与对照无显著差异。几丁寡糖处理后对苎麻株高和原麻产量以120×10-6稀释液处理最高,分别为195.47cm和950.87g,与对照相比分别增加22.88%和25.40%,60×10-6、80×10-6和200×10-6稀释液处理次之,均显著高于对照,其他浓度处理与对照无显著差异。水杨酸处理后对苎麻株高和原麻产量以0.2mmol/L处理最高,分别为183.33cm和921.38g,与对照相比分别增加15.25%和21.51%,0.4、0.8、1.2mmol/L处理次之,均显著高于对照,其余浓度处理与对照无显著差异,3.0mmol/L处理对苎麻生长有抑制作用,其株高和产量均低于对照(图3~4)。
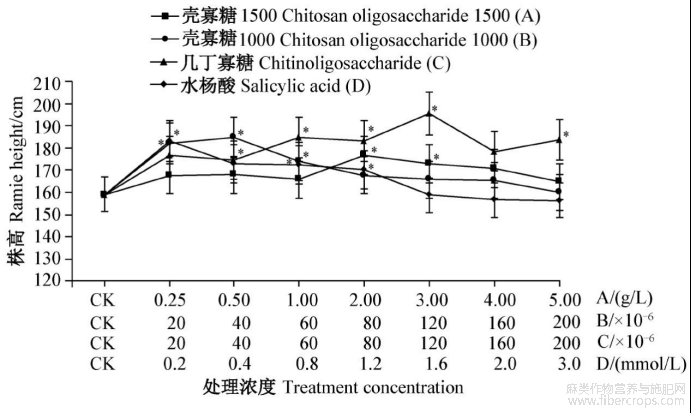
图3 4种诱抗剂不同浓度处理后苎麻的株高
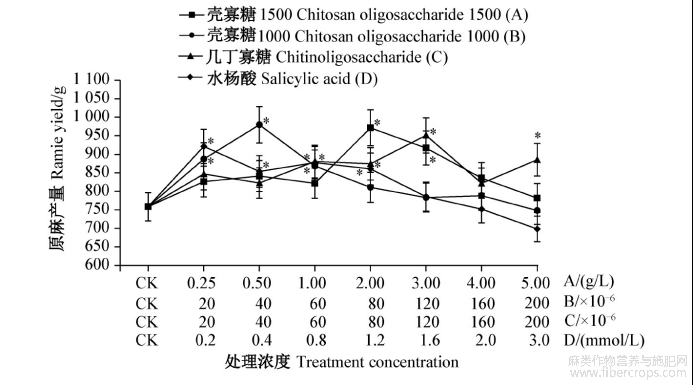
图4 4种诱抗剂不同浓度处理后苎麻的原麻产量
3讨论
本研究结果显示,4种诱抗剂在室内试验和田间试验中的总体趋势基本一致,80×10-6~120×10-6几丁寡糖稀释液能较好地将苎麻夜蛾为害程度指数控制在10左右,其次为80×10-6~120×10-6壳寡糖1000稀释液,能将苎麻夜蛾为害程度指数控制在20左右,而0.8~3.0mmol/L水杨酸和1.0~5.0g/L壳寡糖1500对苎麻的诱抗效果相近,能将苎麻夜蛾为害程度指数控制在30~40之间,但低浓度的水杨酸能刺激苎麻夜蛾的取食。壳寡糖1500、壳寡糖1000和几丁寡糖的室内试验结果和田间试验结果稍有差异,这可能与壳寡糖为2~8个氨基寡糖混合物有关,且不同分子量的寡糖生理活性存在差异。另外,在田间诱抗试验中,诱抗剂对苎麻夜蛾的产卵选择有趋避作用(安晓霞,2014),这可能是造成几丁寡糖处理田间诱抗效果优于室内试验的主要原因。寡糖类诱抗剂诱导植物产生防卫反应信号是多途径的、复杂的,但其中很多信号转导机制尚不明确。据Yama‐daetal.(1993)报道几丁寡糖能诱导植物产生抗毒素,杜昱光等(2002)用壳寡糖处理烟草可不同程度地提高其叶片中过氧化氢酶、多酚氧化酶、过氧化物酶、苯丙氨酸解氨酶和β-1,3葡聚糖酶的活性。外源水杨酸处理花生(鄢洪海等,2006)、香蕉(江新华等,2006)和水稻(王霞等,2007)等植物可以诱导其对多种植物病虫害(苗进等,2007)产生局部和系统抗性。这些被诱导的途径在苎麻中也有可能发生,从而增强苎麻的抗病虫能力。
在田间跟踪测产中,4种诱抗剂在适宜浓度下相较于对照均能显著提高株高和原麻产量,但4种诱抗剂最适浓度处理之间的产量并无显著差异。各诱抗剂适宜浓度分别为2.0~3.0g/L壳寡糖1500、20×10-6~60×10-6壳寡糖1000稀释液、几丁寡糖60×10-6~120×10-6稀释液和0.8~1.2mmol/L水杨酸。综合来看,4种诱抗剂在适宜浓度施用时表现良好,可以诱导苎麻对苎麻夜蛾产生抗性,且能显著促进原麻增产。以提高苎麻产量和对苎麻夜蛾的抗性为评价标准,在田间综合增产和防虫功效优先条件下,推荐施用几丁寡糖80×10-6~120×10-6稀释液和壳寡糖100040×10-6~80×10-6稀释液,其次为0.8~1.2mmol/L水杨酸和2.0~3.0g/L壳寡糖1500。这与安晓霞(2014)所得0.8mmol/L水杨酸及3g/L壳寡糖1500处理能使苎麻产量及株高有所提高的结果一致,且本试验中几丁寡糖和壳寡糖1000的综合效果更优。
刘俊杰等(2020)研究发现使用壳寡糖预处理的拟南芥植株内源水杨酸含量明显升高。在本试验中,壳寡糖处理组与水杨酸处理组总体变化趋势基本吻合,壳寡糖能否诱导苎麻内源水杨酸含量升高还需进一步研究验证。下一步可探讨几丁寡糖和壳寡糖1000处理后苎麻中如多酚氧化酶、过氧化物酶和超氧化物歧化酶等防御酶的活性变化,苎麻相关次生代谢物对苎麻夜蛾的取食和产卵等行为的趋避作用,以及对原麻纤维的支数、强力等特性。
参考文献
[1]An XX.2014.Effects of elicitors on resistance to Cocytodes coerulea Guenée(Lepidoptera:Noctuidae) and yield in ramie.Master thesis.Beijing:Chinese Academy of Agricultural Sciences(in Chinese)[安晓霞.2014.诱抗剂对苎麻抗夜蛾能力及产量的影响.硕士学位论文.北京:中国农业科学院]
[2]Appu M,Muthukrishnan S.2014.Foliar application of salicylic acid stimulates flowering and induce defense related proteins in finger millet plants.Universal Journal of Plant Science,2(1):14-18
[3]Ayoubi N,Soleimani MJ.2015.Possible effects of pathogen inoculation and salicylic acid pre-treatment on the biochemical changes and proline accumulation in green bean.Archives of Phytopathology and Plant Protection,48(3):212-222
[4]Chen JK,Dong GY,Yu CM,Chen P,Gao G,Chen KM,Wang XF,Zhu AG.2020.Evaluation of the silage quality of ramie mixed with rice and/or maize straws.Pratacultural Science,37(3):583-591(in Chinese)[陈继康,董国云,喻春明,陈平,高钢,陈坤梅,王晓飞,朱爱国.2020.苎麻与水稻/玉米秸秆混合青贮饲用价值评价.草业科学,37(3):583-591]
[5]Du YG,Bai XF,Zhao XM,Jiang H.2002.The effect of oligochitosan on the activity of defensive enzyme and zymogram of isoenzyme in tobacco leaves.Chinese Journal of Biological Control,18(2):83-86(in Chinese)[杜昱光,白雪芳,赵小明,姜华.2002.壳寡糖对烟草防御酶活性及同工酶酶谱的影响.中国生物防治,18(2):83-86]
[6]Gong XM,Huang DL,Liu YG,Zeng GM,Wang RZ,Wan J,Zhang C,Cheng M,Qin X,Xue WJ.2017.Stabilized nanoscale zerovalent iron mediated cadmium accumulation and oxidative damage of Boehmeria nivea(L.) Gaudich cultivated in cadmium contaminated sediments.Environmental Science&Technology,51(19):11308-11316
[7]Hussain B,War AR,Sharma HC.2014.Jasmonic and salicylic acidinduced resistance in sorghum against the stem borer Chilo partellus.Phytoparasitica,42(1):99-108
[8]Jiang XH,Xie JH,Wang W,Liang GL.2006.Physiological test on salicylic acid induced resistance of banana vasicular wilt.China Fruits,(6):39-42(in Chinese)[江新华,谢江辉,王尉,梁国鲁.2006.水杨酸诱导香蕉抗枯萎病生理试验.中国果树,(6):39-42]
[9]Jie YC.2007.Specification and data standard of ramie germplasm resources description.Beijing:China Agriculture Press,pp.73(in Chinese)[揭雨成.2007.苎麻种质资源描述规范和数据标准.北京:中国农业出版社,pp.73]
[10]Li SJ,Zhu TH.2013.Biochemical response and induced resistance against anthracnose(Colletotrichum camelliae) of camellia(Camellia pitardii) by chitosan oligosaccharide application.Forest Pathology,3(1):67-76
[11]Liu C,Zhao L,Li XH,Fu ZC,Liu F,Teng Y.2020.Accumulation and transfer of mercury and cadmium in ramie from agricultural soils.Journal of Agro-Environment Science,39(5):1034-1042(in Chinese)[刘冲,赵玲,李秀华,傅赵聪,刘方,滕应.2020.苎麻对农田土壤中汞、镉的吸收累积特征研究.农业环境科学学报,39(5):1034-1042]
[12]Liu JJ,Jia XC,Zhao XM,Wang WX,Yin H.2020.Function of SPINDLY in chitosan oligosaccharide induced resistance to pst DC3000 in Arabidopsis.Acta Botanica Boreali-Occidentalia Sinica,40(5):766-772(in Chinese)[刘俊杰,贾晓晨,赵小明,王文霞,尹恒.2020.SPINDLY在壳寡糖诱导拟南芥抗丁香假单胞菌中的功能.西北植物学报,40(5):766-772]
[13]Liu XL,Lou YG.2018.Comparison of the defense responses in rice induced by brown planthopper Nilaparvata lugens(Stål) and white-backed planthopper Sogatella furcifera(Horváth).Journal of Plant Protection,45(5):971-978(in Chinese)[刘晓丽,娄永根.2018.褐飞虱与白背飞虱为害诱导水稻防御反应的比较.植物保护学报,45(5):971-978]
[14]Ma H.2019.Effects of two inducing agents on growth and disease resistance of tomato seedlings.Master thesis.Shenyang:Shenyang Agricultural University(in Chinese)[马赫.2019.两种诱抗剂对番茄幼苗生长和抗病性的影响.硕士学位论文.沈阳:沈阳农业大学]
[15]Ma JW.1989.Cocytodes coerulea harm elm.Plant Protection,15(4):60(in Chinese)[马继武.1989.苎麻夜蛾为害榆树.植物保护,15(4):60]
[16]Miao J,Li GP,Han BY.2007.Recent developments in effect and mechanism of salicylic acid and methyl salicylate on plant resistance to pests.Chinese Journal of Tropical Crops,28(1):111-114(in Chinese)[苗进,李国平,韩宝瑜.2007.水杨酸和水杨酸甲酯在植物抗虫中的作用及机制研究进展.热带作物学报,28(1):111-114]
[17]Mostafanezhad H,sahebani N,Nourinejhad Zarghani S.2014.Control of root-knot nematode(Meloidogyne javanica) with combination of Arthrobotrys oligospora and salicylic acid and study of some plant defense responses.Biocontrol Science and Technology,24(2):203-215
[18]Qin RJ.2018.Effects of plant inducer combined with controlledrelease urea on nitrogen use efficiency and yield of wheat and corn.Master thesis.Tai’an:Shandong Agricultural University(in Chinese)[秦瑞劼.2018.控释尿素配施植物诱抗剂对小麦玉米周年产量及氮肥利用率的影响.硕士学位论文.泰安:山东农业大学]
[19]Shao SQ,Li JZ.2002.The progress about study of plant elicitors.Pesticides,41(6):12-14(in Chinese)[邵素琴,李建中.2002.植物诱抗剂研究进展.农药,41(6):12-14]
[20]Sun K,Yu YT,Zeng LB.2015.Effect of chitosan oligosaccharide on inducing insect resistance of ramie.Studies on Insects in Central China,(1):307(in Chinese)[孙凯,余永廷,曾粮斌.2015.壳寡糖对苎麻的诱导抗虫作用.华中昆虫研究,(1):307]
[21]Wang HQ,Miao FJ,Li B,Wan H,Liu SX.2013.Selection of the optimal inducers for potato scab.Journal of Southern Agriculture,44(7):1125-1129(in Chinese)[王宏虬,缪福俊,李彪,万红,刘绍雄.2013.马铃薯疮痂病最适诱抗剂筛选试验.南方农业学报,44(7):1125-1129]
[22]Wang X,Du MH,Zhou GX,Cheng JA,Lou YG.2007.Role of salicylic acid and H2O2signaling pathways in the production of rice volatiles induced by the rice brown planthopper Nilaparvata lugens.Journal of Zhejiang University(Agriculture and Life Sciences),33(1):15-23(in Chinese)[王霞,杜孟浩,周国鑫,程家安,娄永根.2007.水杨酸与过氧化氢信号途径在褐飞虱诱导的水稻挥发物释放中的作用.浙江大学学报(农业与生命科学版),33(1):15-23]
[23]Wang YE,Wang SH.1987.Preliminary observation on Cocytodes coerulea.Hubei Agricultural Sciences,26(3):20(in Chinese)[王玉娥,王绍华.1987.苎麻夜蛾的初步观察.湖北农业科学,26(3):20]
[24]Xiong CC,Zeng LB,Li JZ,Wang HW,Xue ZD.2012.Occurrence and control of main diseases on ramie.Plant Fiber Sciences in China,34(5):220-225(in Chinese)[熊常财,曾粮斌,李景柱,汪红武,薛召东.2012.苎麻主要病害的发生及防治.中国麻业科学,34(5):220-225]
[25]Yamada A,Shibuya N,Kodama O,Akatsuka T.1993.Induction of phytoalexin formation in suspension-cultured rice cells by N-acetylchitooligosaccharides.Bioscience,Biotechnology,and Biochemistry,57(3):405-409
[26]Yan HH,Zhao ZQ,Wang Y,Guo SQ,Xia SC,Chi SQ.2006.Change of defense enzymatic activities after treatment with different salicylic acid in peanut.Journal of Peanut Science,35(4):20-22(in Chinese)[鄢洪海,赵志强,王琰,国姝琼,夏淑春,迟胜起.2006.水杨酸处理对花生主要防御酶活性的影响.花生学报,35(4):20-22]
[27]Yu YT,Xue ZD,Zeng LB,Zhang G,Chen Q,Zhu AG.2011.Identification of a nematode isolate from rot root of ramie.Journal of Northwest A&F University(Natural Science Edition),39(7):105-109(in Chinese)[余永廷,薛召东,曾粮斌,张岗,陈权,朱爱国.2011.1种苎麻根腐病线虫的鉴定.西北农林科技大学学报(自然科学版),39(7):105-109]
[28]Zeng LB,Shen AR,Chen J,Yan Z,Liu TM,Xue ZD,Yu YT.2016.Transcriptome analysis of ramie(Boehmeria nivea L.Gaud.) in response to ramie moth(Cocytodes coerulea Guenée) infestation.BioMed Research International,2016:1-10
[29]Zeng LB,Xue ZD,Yan ZY,Yu YT,Yang RL.2011.Relationship between main chemical compositions in leaves of ramie and resistance to Cocytodes coerulea.Hunan Agricultural Sciences,(19):77-78(in Chinese)[曾粮斌,薛召东,严智燕,余永廷,杨瑞林.2011.苎麻叶片主要化学成分与抗苎麻夜蛾关系的研究.湖南农业科学,(19):77-78]
[30]Zeng LB,Xue ZD,Yu YT,Yan ZY,Yang RL.2013.The regularity and control technology of Cocytodes coerula.Hunan Agricultural Sciences,(10):23-24(in Chinese)[曾粮斌,薛召东,余永廷,严智燕,杨瑞林.2013.苎麻夜蛾发生规律变化及其防治技术.湖南农业科学,(10):23-24]
文章摘自:翟杨,罗晨,牛艳斌,汪洪鹰,杨媛茹,薛召东,陈佳,曾粮斌.四种诱抗剂对苎麻生长发育和抗苎麻夜蛾特性的影响[J].植物保护学报,2022,49(03):809-815.
