摘 要:本试验以湘苎7号作为材料,探究6-苄氨基腺嘌呤(6-benzyladenine,6-BA)调控增强苎麻抗旱性的生理机制,结果表明,不同浓度6-BA处理苎麻叶片脯氨酸(PRO)含量、可溶性糖(SS)和可溶性蛋白(SP)含量的积累量随着处理天数的增加整体呈现不断上升的趋势,株高、茎粗、叶面积、生物量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性、抗坏血酸过氧化物酶(APX)活性均表现出先升高后下降的趋势,多以干旱胁迫24d是最高,而丙二醛(MDA)含量、叶片相对电导率与W0相比逐渐降低,相对含水量呈持续下降。通过对苎麻叶片各项生理生化指标进行综合分析得出,不同浓度6-BA处理抗旱性综合排序为W2(100mgL−1)>W3(150mgL−1)>W1(50mgL−1)>W4(mgL−1),由此可得,100mgL−16-BA处理下干旱胁迫后苎麻的缓解效应最好。
关键词:6-BA;苎麻;生理特性;干旱
苎麻(Boehmeria nivea L.),是我国传统的经济作物,其纤维品质优质,在国内外市场享有名誉[1],主产地来自中国西南部的山坡、丘陵一带[2]。目前,中国是苎麻最大的生产国家,2024年我国苎麻种植面积2.3万公顷左右,年均原麻产量达5.5万吨,均远超其他国家[3]。苎麻纺织品成为了我国重要的出口商品,麻纺行业的快速发展助力了我国经济进一步提高[4]。苎麻功能用途不断被发掘,主要表现在饲用喂养、麻地膜、保水固土、修复重金属污染以及改善土壤理化性质等方面[5]。
近些年,全球农作物受干旱天气影响面积日益扩大[6]。干旱引起植株叶片萎蔫相对含水量降低,从而减缓植物生长发育[7]。长时间干旱限制了农作物生长,导致作物产量和品质严重下降[8]。湖南地区夏季易出现高温干旱气候,淡水资源分布不均,降雨量不充分,年平均降水量差异大,这些不利因素对苎麻栽培产生了较大的影响[9]。因此,有效解决苎麻种植过程中面临的干旱问题对我国苎麻高产栽培具有重大的意义。
6-苄氨基腺嘌呤(6-benzyladenine,6-BA)是一种促进植物生长的激素物质,它可以维持逆境胁迫下植物地上部与地下部之间矿质元素的平衡,提高植物抗逆能力[10]。杨喆等[11]研究表明,水稻分蘖期喷施25mgL−16-BA溶液,短期内会对水稻有较好的抵抗干旱胁迫作用,暂时抑制细胞内氧化物的产生,提高细胞防御水平,有效缓解干旱胁迫对水稻生理代谢功能的损伤。李彩龙等[12]研究表明,干旱胁迫能显著抑制苹果幼苗的生长,喷施50mgL−16-BA可以减增强抗氧化酶活性,积累渗透调节物质,提高生物膜稳定性来缓解干旱胁迫对植株的伤害。王军等[13]研究表明,烟草旺长期喷施200μmolL−16BA,能有效提高烤烟的耐旱性。现有研究表明,外源喷施适宜浓度的6-BA能够有效缓解水分[14]、高温[15]、重金属[16]等多种逆境胁迫,但关于提高苎麻抗旱性方面的研究还未见报道。本研究采用盆栽试验法,选取湘苎7号为试验材料,探究6-BA调控增强苎麻抗旱性的生理机制,以期为苎麻抗旱性研究和栽培生产提供科学的理论支持。
1 材料与方法
1.1 试验材料
试验材料为湖南地区主栽苎麻品种湘苎7号,由湖南农业大学苎麻研究所提供。
1.2 试验设计
本试验采取盆栽试验法,共设置6个处理组,分别为CK(正常供水组)、W0(干旱胁迫组)、W1(干旱+50 mg L−16-BA)、W2(干旱+100 mg L−1 6-BA)、W3(干旱+150 mg L−16-BA)、W4(干旱+200 mg L−16-BA),每个处理重复9次,总共54盆,每盆种植2株。每日早晚向苎麻叶片正反两面喷施200mL不同浓度的6-BA处理液,共连续进行3 d,以叶片表面湿润且无明显水珠滴下为标准。第4天对除正常供水组以外的盆栽进行干旱胁迫处理,利用土壤水分测定仪和称重法对土壤含水量进行严格管控,并于每天下午将水分补充至胁迫区间。干旱开始后第12天、24天和36天,采集成熟苎麻叶片进行各项生理生化指标的测定。
1.3 测定项目及方法
1.3.1 苎麻农艺性状的测定
株高:在每组处理中将随机挑选5株苎麻,利用直尺对植株基部至顶端的高度进行测量,并计算所得数据的平均值。茎粗:在株高测量完成后,对同一株苎麻采用游标卡尺测量其中部的茎秆粗度,并计算所得数据的平均值。叶面积:用卷尺测量苎麻植株倒三叶的叶片长度和叶片宽度,根据公式算出每片叶的均值,叶面积=长×宽×0.7。地上部与地下部鲜干重:在第36天处理结束时,从中挑取3株生长基本一致的植株,使用电子秤准确称量苎麻地上及地下部的鲜物质重量,在90℃的烘箱中对植株进行杀青15 min处理,接着再75℃烘干至苎麻恒重,利用电子秤称量其干物质重量。
1.3.2 膜脂过氧化的测定
采用烘干法测定[17]叶片相对含水量;采用电导仪测定[17]叶片相对电导率;参考硫代巴比妥酸法测定[17]丙二醛(Malondialdehyde,MDA)含量。
1.3.3 渗透调节物质含量的测定
采用酸性茚三酮比色法[17]测定脯氨酸(Proline,Pro)含量、采用蒽酮乙酸乙酯法[17]测定可溶性糖(Soluble Sugar,SS)含量、采用考马斯亮蓝G-250染色法[17]测定可溶性蛋白(Soluble protein,SP)含量,均采用上海茁彩生物科技公司试剂盒进行测定[17]。
1.3.4 抗氧化酶活性的测定
参考愈创木酚法测定[18]过氧化物酶(Peroxidase,POD)活性;参考黄嘌呤氧化酶法测定[18]超氧化物歧化酶(Superoxide dismutase,SOD)活性;参考紫外吸收法法测定[18]过氧化氢酶(Catalase,CAT)活性;依据茁彩生物科技公司试剂盒进行测定[18]抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)活性。
1.4 数据处理与分析
使用Microsoft Excel 2019对试验数据整理与作图,采用SPSS 26.0进行数据相关性分析,Duncan法进行多重比较检验,显著水平均为P<0.05。
2 结果与分析
2.1 6-BA 对干旱胁迫下苎麻农艺性状的影响
2.1.1 6-BA 对干旱胁迫下苎麻株高的影响
由表1可知,干旱胁迫对湘苎7号株高产生抑制作用,且随着胁迫天数的延长,抑制程度不断增加。胁迫开始后第12天、24天和36天,湘苎7号株高在W0处理下较CK分别降低19.80%、23.70%和27.55%。第12天时,株高在W1、W2、W3和W4处理下,相比于W0处理分别提高3.84%、7.25%、5.55%和3.21%;第24天时,与W0处理相比,W1、W2、W3和W4处理下苎麻株高分别提高4.36%、10.91%、6.85%和6.12%;第36天时,与W0处理相比,W1、W2、W3和W4处理下的株高分别提高12.60%、17.26%、14.06%和11.58%,且各处理组均与W0处理存在显著性差异(P<0.05)。
表1 干旱胁迫下不同浓度6-BA处理对湘苎7号株高的影响
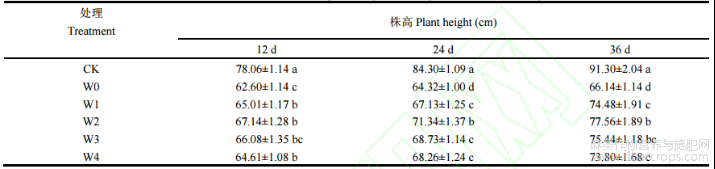
注:同列数值不同字母表示在5%概率水平差异显著。CK:正常供水组;W0:干旱胁迫组:W1:干旱+50mg L−1 6-BA;W2:干旱+100mg L−1 6-BA;W3:干旱+150mg L−1 6-BA;W4:干旱+200mg L−1 6-BA。
2.1.2 6-BA 对干旱胁迫下苎麻茎粗的影响
胁迫开始后第12天、24天和36天,湘苎7号茎粗在W0处理下较CK分别降低7.91%、13.48%和16.99%(表2)。植株茎粗随着外源喷施6-BA浓度的增加,呈现出先升高后降低的变化规律。第12天时,湘苎7号品种茎粗在W1、W2、W3和W4处理下,相比于W0处理分别提高2.04%、2.86%、2.31%和1.90%;第24天时,与W0处理相比,W1、W2、W3和W4处理下植株茎粗分别提高3.09%、4.97%、4.30%和1.07%;第36天时,与W0处理相比,W1、W2、W3和W4处理下的茎粗分别提高3.45%、8.77%、5.05%和4.25%,且W2处理对干旱胁迫下湘苎7号茎粗的促进效果最佳。
表2 干旱胁迫下不同浓度6-BA处理对湘苎7号茎粗的影响
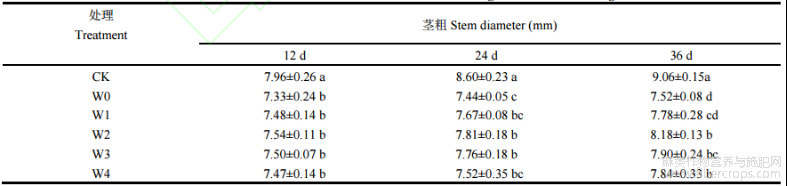
同列数值不同字母表示在5%概率水平差异显著。处理同表1。
2.1.3 6-BA对干旱胁迫下苎麻叶面积的影响
外源喷施不同浓度的6-BA后,植株叶面积得到一定程度的缓解,主要表现为“低促高抑”的变化规律(表3)。胁迫开始后第12天、24天和36天,湘苎7号叶面积在W0处理下与CK相比分别降低18.61%、21.02%和25.84%。第12天时,湘苎7号品种叶面积在W1、W2、W3和W4处理下,相比于W0处理分别提高8.98%、13.34%、6.75%和3.41%;第24天时,与W0处理相比,W1、W2、W3和W4处理下叶面积分别提高7.87%、17.24%、10.56%和9.05%;胁迫第36天时,与W0处理相比,W1、W2、W3和W4处理下苎麻叶面积分别提高11.24%、21.60%、14.38%和11.09%。
表3 干旱胁迫下不同浓度6-BA处理对湘苎7号叶面积的影响
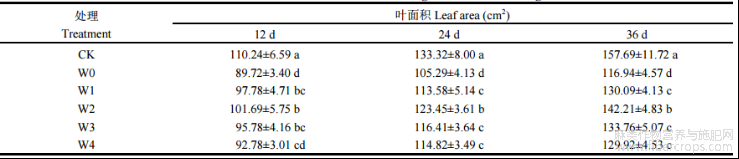
同列数值不同字母表示在 5%概率水平差异显著。处理同表 1。
2.1.4 6-BA对干旱胁迫下苎麻地上与地下部鲜干重的影响
湘苎7号地上部鲜重及干重在W0处理下相较于CK分别下降29.15%和40.52%(表4)。湘苎7号在W1、W2、W3和W4处理下,与W0处理相比,植株地上部鲜重分别提高7.42%、14.50%、11.94%和4.56%,地上部干重分别提高23.61%、42.84%、30.21%和19.51%,W2处理的苎麻地上部产量提升最多。
在W0处理下,湘苎7号地下部鲜重及干重相较于CK分别下降35.70%和27.51%。湘苎7号在W1、W2、W3和W4处理下,与W0处理相比,苎麻地下部鲜重分别提高14.21%、18.75%、15.90%和6.50%,地下部干重分别提高12.78%、25.64%、14.50%和6.98%。
表4 干旱胁迫下不同浓度6-BA处理对湘苎7号生物量的影响
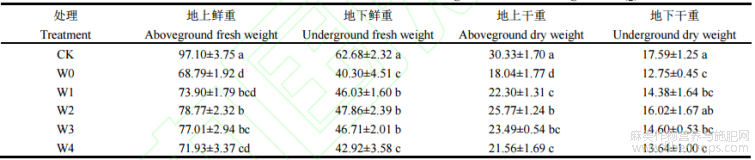
同列数值不同字母表示在5%概率水平差异显著。处理同表1。
2.2 6-BA 对干旱胁迫下苎麻叶片膜脂过氧化的影响
2.2.1 对苎麻丙二醛含量的影响
由图1可知,随着干旱天数的不断延长,苎麻叶片丙二醛含量呈现逐渐上升的趋势,并在第36天时达到最大值。经过不同浓度的6-BA处理后,各时期叶片丙二醛含量均低于W0处理组。胁迫第36天,苎麻叶片丙二醛含量在W1、W2、W3和W4处理下,与W0处理相比分别降低10.52%、25.56%、13.84%和7.69%,W2处理的丙二醛含量降幅最大,与W0处理之间达到显著性差异水平(P<0.05)。可见,在干旱环境下喷施适宜浓度的6-BA能够降低苎麻丙二醛含量,缓解干旱逆境下膜脂过氧化作用对植物造成的伤害。
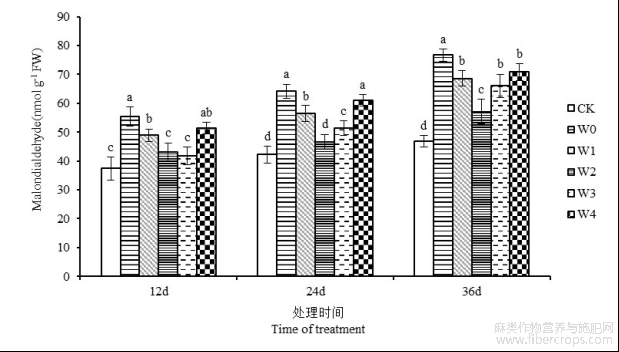
图1 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片丙二醛含量的影响
2.2.2 对苎麻相对电导率的影响
叶片相对电导率与植物细胞膜损伤程度密切相关。由图2可知,随着处理天数的增加,苎麻叶片相对电导率总体表现为持续上升的变化趋势。在胁迫后期第36天,叶片相对电导率达到最大值,较CK对照组提高40.75%。叶面喷施6-BA可以有效抑制苎麻遭遇干旱胁迫时相对电导率的升高。除干旱胁迫第12天外,其余时间段苎麻叶片相对电导率均在W2处理下降低幅度最大,比W0处理组分别降低24.56%和25.13%,且干旱处理组存在显著性差异(P<0.05)。表明6-BA处理可以减轻干旱胁迫造成的植物体内电解质外渗,从而能够有效缓解苎麻细胞膜结构受损。
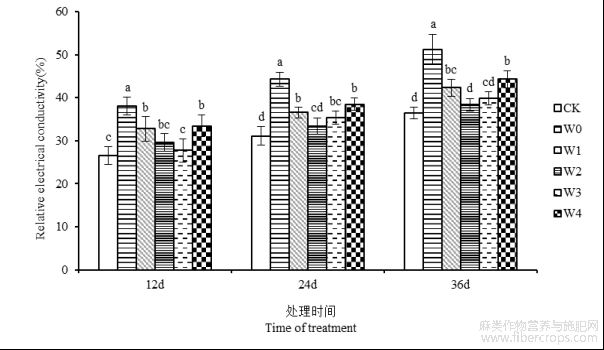
图2 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片相对电导率的影响
2.2.3 对苎麻相对含水量的影响
由图3可知,在干旱条件下,随胁迫处理时间的延长,苎麻叶片相对含水量呈持续下降的趋势。第36天,苎麻叶片相对含水量在W0处理下较CK对照组降低25.65%。喷施不同浓度的6-BA对干旱胁迫下苎麻叶片相对含水量产生促进作用。干旱胁迫第36天时,苎麻叶片相对含水量在W1、W2、W3和W4处理下,与W0处理相比分别提高9.66%、16.15%、10.62%和4.07%,并且以W2处理的促进效果最佳。说明喷施6-BA有利于干旱胁迫下苎麻的水分供应,该浓度水平对苎麻叶片相对含水量具有较好的促进效应。

图3 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片相对含水量的影响
2.3 6-BA 对干旱胁迫下苎麻叶片渗透调节物质的影响
2.3.1 对苎麻可溶性糖含量的影响
由图4可知,干旱胁迫下苎麻叶片可溶性糖含量呈上升的趋势。第12、24和36天,苎麻叶片可溶性糖含量在干旱处理组较CK对照组分别提高4.88%、10.51%和12.96%。干旱胁迫下喷施6-BA能够进一步提高苎麻叶片中可溶性糖含量。胁迫第36天时,苎麻叶片可溶性糖含量在W1、W2、W3和W4处理下,与W0处理相比分别提高16.54%、23.68%、22.33%和11.89%,W2处理的叶片可溶性糖含量达到最高值,为11.96mg g−1。由此可见,干旱胁迫下喷施6-BA有利于苎麻叶片中可溶性糖的积累,从而提高植物保水能力,缓解干旱逆境对植物膜系统的伤害。
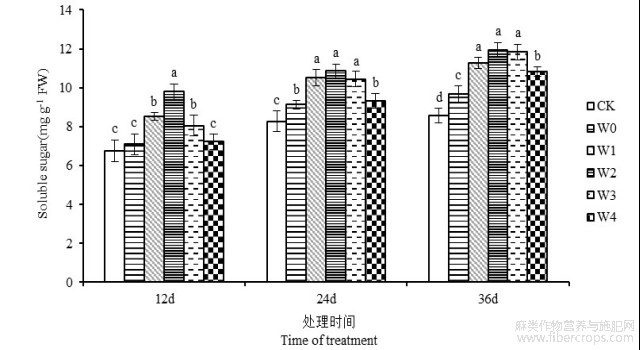
图4 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片可溶性糖含量的影响
2.3.2 对苎麻可溶性蛋白含量的影响
湘苎7号各处理的叶片可溶性蛋白含量的变化情况如图5所示。由图可知,苎麻叶片可溶性蛋白含量随胁迫时间的延长呈上升趋势,并在第36天时达到峰值,较CK对照提高34.95%。干旱胁迫下6-BA处理的苎麻叶片可溶性蛋白含量有所提高。胁迫第36天,苎麻叶片可溶性蛋白含量在W1、W2、W3和W4处理下,与W0处理相比分别提高18.70%、26.97%、20.50%和15.82%,各浓度水平均与干旱胁迫组达到差异显著(P<0.05)。可见,喷施6BA能够促进干旱胁迫下苎麻叶片中可溶性蛋白的产生,从而提高苎麻抗旱性。
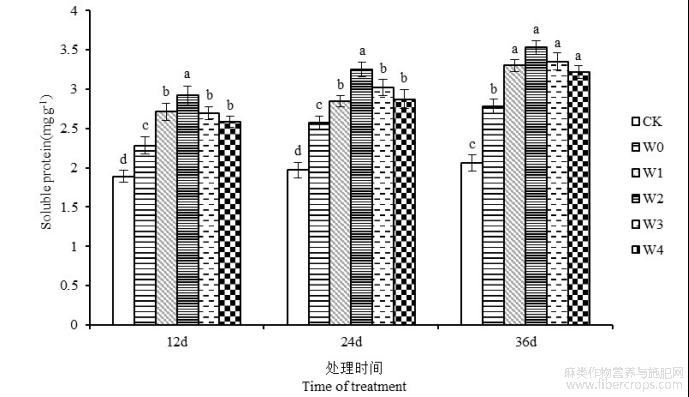
图5 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片可溶性蛋白含量的影响
2.3.3 对苎麻脯氨酸含量的影响
由图6可知,干旱胁迫导致苎麻叶片脯氨酸含量显著提高(P<0.05)。胁迫开始后第12、24和36天,湘苎7号叶片脯氨酸含量在W0处理下较CK对照组分别提高24.89%、63.90%、90.87%,并在第36天达到峰值。表明干旱胁迫会诱导植物产生大量的脯氨酸,且随着胁迫天数的增长,其促进作用越明显。干旱胁迫第36天,苎麻叶片脯氨酸含量在W1、W2、W3和W4处理下,与W0处理相比分别提高36.78%、46.92%、40.72%和35.42%,W2处理下的苎麻叶片中脯氨酸含量达最大值,为280.97 mg g−1。可见,外源喷施6-BA能够有效提高干旱胁迫下苎麻叶片脯氨酸含量,以应对水分亏缺状态。
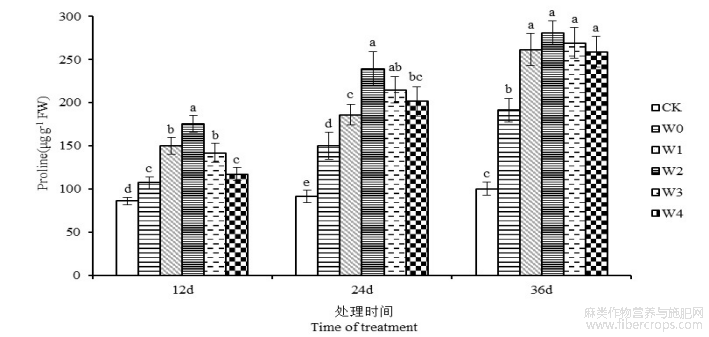
图6 干旱胁迫不同浓度6-BA处理对湘苎7号叶片脯氨酸含量的影响
2.4 6-BA 对干旱胁迫下苎麻叶片抗氧化酶活性的影响
2.4.1 对苎麻超氧化物歧化酶活性的影响
由图7可知,随着胁迫天数的增加,苎麻叶片SOD活性总体表现为先上升后下降的趋势。胁迫开始后第12、24和36天,苎麻SOD活性在W0处理下较CK分别提高了19.58%、26.94%、23.63%。干旱胁迫下喷施一定浓度的6-BA对苎麻叶片中的SOD活性具有提升作用,并且均高于W0处理。胁迫第24天时,苎麻叶片SOD活性在W1、W2、W3和W4处理下,与W0处理相比分别提高19.43%、29.18%、24.42%和10.96%,W2处理的叶片SOD活性达到峰值,为521.70 U g−1。说明外源喷施适宜浓度的6-BA能够提高苎麻叶片SOD活性,增强抗氧化防御机制,从而降低干旱胁迫对植株造成的损伤。
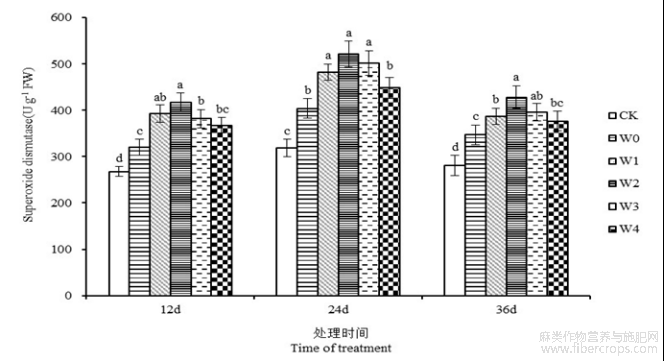
图7 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片SOD活性的影响
2.4.2 对苎麻过氧化物酶活性的影响
由图8可知,干旱胁迫导致湘苎7号叶片POD活性有所提高。第12、24和36天,湘苎7号叶片POD活性在W0处理下较CK分别提高13.71%、18.06%、14.52%。干旱胁迫下外源喷施6-BA能够进一步促进叶片中POD活性的提高,且各浓度处理的POD活性均高于干旱处理组。胁迫开始后第24天,苎麻叶片POD活性在W1、W2、W3和W4处理下,相较于W0处理分别提高了18.99%、25.59%、14.24%和12.66%,W2处理下叶片POD活性高达777.46 U g−1,为最大值,且与W0处理达到显著性差异(P<0.05)。表明,叶面喷施6-BA能够提高苎麻叶片中POD活性,增强抗氧化系统,抵御干旱逆境对植株的损伤。
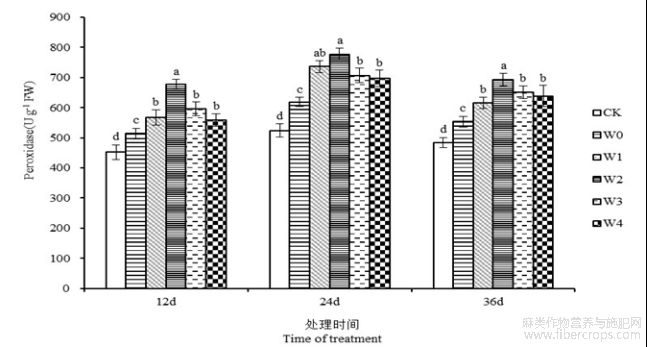
图8 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片POD活性的影响
2.4.3 对苎麻过氧化氢酶活性的影响
由图9可知,随着干旱时间的增加,各处理的苎麻叶片CAT活性呈现先升高后降低的趋势。第12、24和36天,湘苎7号叶片CAT活性在W0处理下较CK分别提高21.88%、26.30%、24.37%,并在第24天达到峰值。干旱胁迫下喷施不同浓度的6-BA后,苎麻叶片CAT活性均高于W0处理。干旱胁迫第24天,苎麻叶片CAT活性在W1、W2、W3和W4处理下,与W0处理相比分别提高19.77%、27.74%、21.88%和13.27%,W2处理的促进效果最佳,CAT活性为1233.13 U g−1,且与W0处理之间存在显著性差异(P<0.05)。表明,干旱胁迫会导致苎麻叶片中CAT活性的提高,而叶面喷施6-BA有利于植株CAT活性进一步提升。
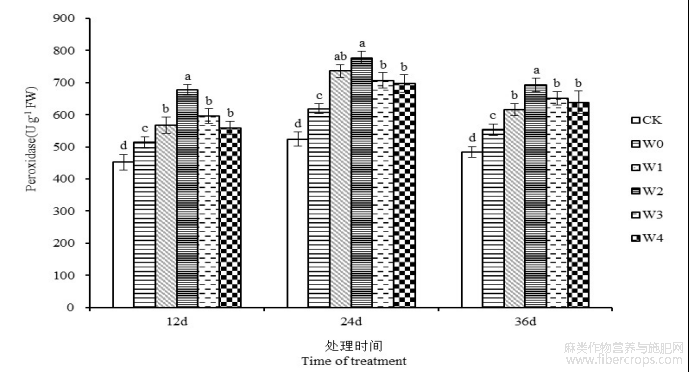
图9 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片CAT活性的影响
2.4.4 对苎麻抗坏血酸过氧化物酶活性的影响
图10是干旱胁迫下叶面喷施6-BA对苎麻叶片APX活性的影响情况。第12、24和36天,苎麻叶片APX活性在W0处理下较CK分别提高21.45%、25.24%、22.37%。干旱胁迫下施加一定浓度的6-BA能够有效提高苎麻叶片APX活性。干旱胁迫第24天,苎麻叶片APX活性在W1、W3和W4处理下,与W0处理相比分别提高25.80%、31.81%和12.08%,W2处理下叶片APX活性达到最大值,为2.28 U g−1,较W0处理提高38.85%,且与各处理组之间存在差异显著性(P<0.05)。
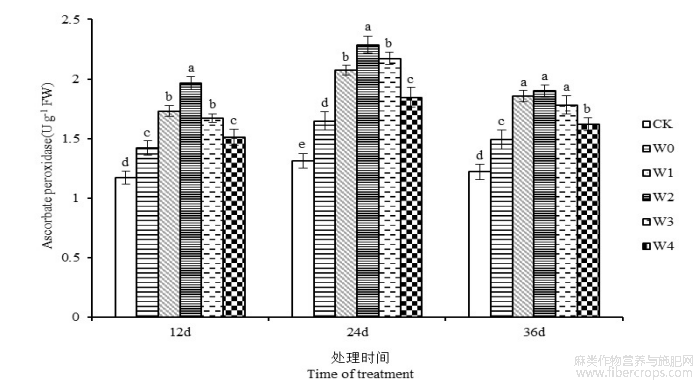
图10 干旱胁迫下不同浓度6-BA处理对湘苎7号叶片APX活性的影响
2.5 干旱胁迫下不同浓度6-BA处理对苎麻抗旱能力的综合评价
运用SPSS26.0软件进行主成分分析。通过计算不同浓度6-BA处理下苎麻各生理生化指标的贡献值,对不同处理的抗旱性进行综合得分排序,得其抗旱能力大小。由表5可知,2个主成分的特征值分别为6.198和2.398均大于1。第一主成分贡献率为61.997%,第二主成分贡献率为23.982%,其方差贡献率达到85.959%,大于85%,具有较好代表性,与6-BA对苎麻干旱胁迫后的缓解效应具有直接联系,可作为评价6-BA对苎麻干旱胁迫后缓解效应的综合指标。第一成分主要综合了APX、CAT、POD、SOD、SP等指标,第二成分主要综合了MDA、相对电导率、相对含水量等指标(表6)。由表7可知,苎麻在各处理下的综合得分排名为W2>W3>W1>W4>W0>CK,由此可得,W2处理下干旱胁迫后苎麻的缓解效应最好。
表5 方差贡献率载荷矩阵
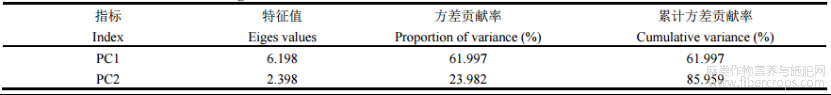
表6 主成分分析后的旋转载荷矩阵
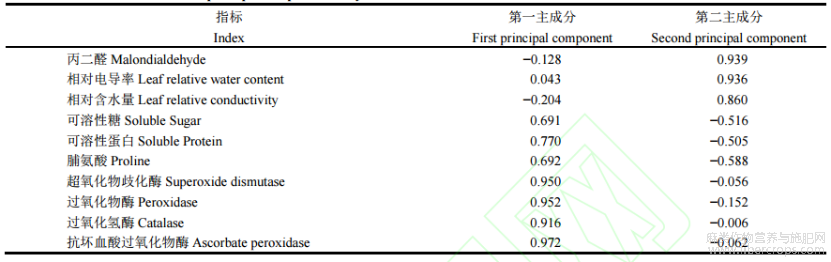
表7 综合得分排名
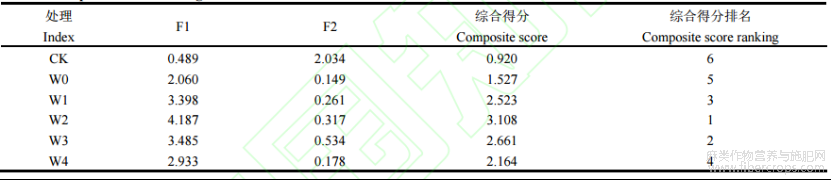
3 讨论
干旱是农业生产中最常见的自然灾害,干旱胁迫会抑制植物的吸收和积累功能。研究发现,在水分亏缺状态下苎麻株高、茎粗和鲜皮重量均显著下降,最终影响植株生物产量的积累[19]。适宜的浓度6-BA有利于提高植株叶片相对含水量,延缓植物细胞衰老,有效减轻干旱胁迫对植物的损伤。李林宇等[20]的研究表明,干旱胁迫下外源喷施6-BA越橘各项生长指标有所缓解,处理浓度为150mg L−1时效果最佳。王金强[21]的研究表明,干旱胁迫条件下,甘薯植株的生长发育过程受明显抑制,主要表现为地上及地下部生物量显著下降,经过不同浓度6-BA处理后,能够有效缓解干旱胁迫对植物的抑制作用,促进植株正常生长。本研究结果表明,干旱胁迫导致湘苎7号的株高、茎粗和叶面积均显著降低,且湘苎7号在干旱胁迫下保持较好的形态特征,这可能是由于湘苎7号具有较好耐旱性。随着外源物质浓度的提高,植株叶面积总体表现为先升高后下降的趋势,且各浓度处理均高于干旱胁迫组。湘苎7号在100mg L−1的6-BA处理下,叶面积、地上部鲜重相较于干旱胁迫组提升显著。这表明喷施适宜浓度的6-BA可以促进干旱胁迫下苎麻的发育生长,有助于植物体对营养元素进行吸收与运送,维持细胞结构稳定,从而促使植物进行正常的生理代谢活动,缓解干旱胁迫对植株生长发育的抑制作用。
植物遭受干旱胁迫时会引起体内活性氧过度积累,导致植物细胞膜的过氧化作用,对植物造成伤害[22-23]。丙二醛是植物过氧化作用的产物,是揭示膜脂过氧化程度的重要指标[24]。相对电导率是反映细胞膜在逆境胁迫下的变化状况的重要生理指标[25-26]。本研究结果发现,干旱胁迫促使植物体内生成了大量的活性氧自由基,随着胁迫时间的延长,苎麻的MDA含量和叶片相对电导率均呈现持续上升的趋势。喷施100mg L-1的6-BA处理能够显著降低苎麻的MDA含量和相对电导率,维持膜细胞稳定性,有效抑制活性氧自由基的产生,从而提高植株抗旱性。这与杨喆[11]等通过外源喷施6-BA缓解干旱胁迫下水稻生长发育的研究结果相同。本试验中,湘苎7号在干旱胁迫下叶片相对含水量均显著下降,而外源喷施6-BA能使苎麻的叶片相对含水量维持在较高水平,说明外源喷施6-BA有利于干旱胁迫下苎麻的水分供应。与胡哲森等[27]针对锥栗幼苗的研究结果相似。渗透调节是植物适应外界环境变化所形成的一种生理机制,通过自身渗透调节来维持细胞具有较高吸水性,保护叶片内的细胞膜不被破坏,促进植物正常生长发育[28-29]。本试验结果表明,随着处理天数的增加,湘苎7号叶片脯氨酸、可溶性糖和可溶性蛋白的积累量均表现出不断上升的趋势。外源喷施6-BA能够进一步提高苎麻体内渗透调节物质的含量,而且在浓度为100 mg L−1的处理下相较于干旱胁迫组显著提升。这与徐封丰等[30]对干旱胁迫下6-BA调控半夏植株渗透调节物质的变化情况相吻合。王军等[13]也有同样的发现,在干旱胁迫下,喷施适宜浓度的6-BA能够显著提高烟草叶片中的可溶性蛋白含量。这些结论进一步证实了6-BA具备稳定剂的功能,能够抑制核酸和蛋白质的分解,维持细胞结构的完整,并促进植物体内渗透物质的积累,进而增强植株抗旱性。
在干旱胁迫条件下,植物通过激活抗氧化防御系统,提升其抗氧化活性,有效地清除因逆境胁迫引发的活性氧自由基,维持细胞内部自由基代谢平衡,从而保护细胞结构的完整性,减少水分亏缺带来的伤害[31-32]。SOD、POD、CAT和APX等抗氧化酶物质已被广泛应用于植物抗逆性研究[33]。研究表明,干旱胁迫会导致玉米、小麦、大豆等多种植物的抗氧化酶活性增加,而这种活性上升与植物的抗旱能力存在一定联系[34-35]。本研究发现,随着处理时间的增加,苎麻各处理的叶片SOD、POD、CAT和APX活性均表现出先升高后下降的趋势,并在胁迫第24天时达到最大值,这说明苎麻能够通过调节抗氧化酶活性来适应环境压力。本研究表明干旱胁迫会导致苎麻叶片SOD、CAT、POD和APX含量显著提升,而外源喷施6-BA能够进一步增加苎麻抗氧化酶活性,以此减轻由活性氧引起氧化损伤,增强植株抗旱性。赵九洲等[36]研究发现,适宜浓度的6-BA能够提高大花蕙兰抗氧化酶活性,随着喷施浓度的增加,保护酶活性呈现先升高后降低的趋势。梁颖等[37]通过PEG模拟干旱胁迫进行试验,探讨了植物生长调节剂6-BA与水稻幼苗抗早性的关系,研究结果显示,在干旱环境下水稻幼苗施加6-BA处理的叶片SOD和POD活性显著提高,说明6-BA一定程度上能够缓解干旱逆境对水稻植株的损伤。
目前,关于6-BA缓解苎麻干旱胁迫的研究并不多见,本研究通过分析外源喷施不同浓度的6-BA对干旱胁迫下苎麻农艺性状和生理特性的影响,揭示了6-BA能够有效减轻干旱逆境对苎麻造成的伤害。据此,本文筛选出了6-BA缓解干旱胁迫下苎麻生长发育的最适浓度,这一发现有望为解决苎麻栽培中遇到的干旱问题提供重要的理论依据和实践指导。不足是苎麻受到干旱胁迫后,常常表现出植株矮化的现象,从而使得纤维很难收获,从而无法判断6-BA处理对干旱胁迫下苎麻纤维产量及品质的影响。同时,研究中所选取的苎麻品种数量有限,这在品种多样性方面存在不足,未来应考虑扩大品种范围。
参考文献
[1]白玉超,李雪玲,黄敏升,孙敬钊,张小龙,曹诣,崔国贤.50年来中国苎麻种植情况与前景展望.作物研究,2014,28:547–550.Bai Y C,Li X L,Huang M S,Sun J Z,Zhang X L,Cao Y,Cui G X.Cultivation of ramie in China over the past 50 years and prospects.Crop Res,2014,28:547–550(in Chinese with English abstract).
[2]王继龙,刘婕仪,刘皖慧,王昕慧,崔丹丹,苏小惠,李林林,佘玮,杨瑞芳,崔国贤.苎麻响应水分胁迫研究进展.中国麻业科学,2020,42(4):178–186.Wang J L,Liu J Y,Liu W H,Wang X H,Cui D D,Su X H,Li L L,She W,Yang R F,Cui G X.Progress of ramie in response to water stress.Plant Fiber Sci China,2020,42(4):178–186(in Chinese with English abstract).
[3]曾业,朱爱国.中国苎麻产业发展迎来“新风口”.(2024-09-02)[2024-09-05].https://www.thecover.cn/.Zeng Y,Zhu A G.Chief Scientist of National Hemp Industry Technology System:the Development of Ramie Industry in China Ushered in a‘New Wind Mouth’.(2024-09-02)[2024-09-05].Cover News.https://www.thecover.cn/.
[4]蒋敏,熊海鹰,张曦.湖南苎麻产业质量情况分.中国纤检,2012,(5):28–31.Jiang M,Xiong H Y,Zhang X.Analysis of the quality of ramie industry in Hunan.China Fiber Inspect,2012,(5):28–31(in Chinese with English abstract).
[5]杨洁,唐昀,申香英,唐文峰,肖群锋.我国苎麻产业现状与振兴发展.中国麻业科学,2022,44(4):253–256.Yang J,Tang Y,Shen X Y,Tang W F,Xiao Q F.Current situation and revitalization of ramie industry in China.Plant Fiber Sci China,2022,44(4):253–256(in Chinese with English abstract).
[6] Yu C Q,Huang X,Chen H,Huang G R,Ni S Q,Wright J S,Hall J,Ciais P,Zhang J,Xiao Y C,Sun Z L,Wang X H,Yu L.Assessing the impacts of extreme agricultural droughts in China under climate and socioeconomic changes.Earth’s Future,2018,6:689–703.
[7] Mukarram M,Choudhary S,Kurjak D,Petek A,Khan M M A.Drought:Sensing,signalling,effects and tolerance in higher plants.Physiol Plant,2021,172:1291–1300.
[8]王春乙,娄秀荣,王建林.中国农业气象灾害对作物产量的影响.自然灾害学报,2007,(5):37–43.Wang C Y,Lou X R,Wang J L.Impacts of agrometeorological disasters on crop yields in China.J Nat Disast,2007,(5):37–43(in Chinese with English abstract).
[9]郝立生,马宁,何丽烨.2022年长江中下游夏季异常干旱高温事件之环流异常特征.干旱气象,2022,40:721–732.Hao L S,Ma N,He L Y.Characteristics of circulation anomalies during the summer anomalous drought and high temperature event in the middle and lower reaches of the Yangtze River in 2022.J Arid Meteorol,2022,40:721–732(in Chinese with English abstract).
[10]Hu J,Ren B Z,Dong S T,Liu P,Zhao B,Zhang J W.6-Benzyladenine increasing subsequent waterlogging-induced waterlogging tolerance of summer maize by increasing hormone signal transduction.Annals New York Acad Sci,2021,1509:89–112.
[11]杨喆,唐才宝,钱婧雅,周伟江,陈光辉,王悦.外源6-BA和BR对干旱胁迫下水稻分蘖期光合色素含量及抗氧化系统的影响.分子植物育种,2021,19:2733–2739.Yang Z,Tang C B,Qian J Y,Zhou W J,Chen G H,Wang Y.Effects of exogenous 6-BA and BR on photosynthetic pigment content and antioxidant system at tillering stage of rice under drought stress.Mol Plant Breed,2021,19:2733–2739(in Chinese with English abstract).
[12]李彩龙,李毛毛,高彦龙,张仲兴,张德,王双成,缐旭林,王延秀.外源6-BA对干旱胁迫下苹果砧木M26的生理效应.甘肃农业大学学报,2022,57(5):126–137.Li C L,Li M M,Gao Y L,Zhang Z X,Zhang D,Wang S C,Beng X L,Wang Y X.Physiological effects of exogenous 6-BA on apple rootstock M26 under drought stress.J Gansu Agric Univ,2022,57(5):126–137(in Chinese with English abstract).
[13]王军,陈帆,温明霞,李小龙,饶兴义,夏志林,许本波.6-BA处理对烤烟耐旱性的影响.作物研究,2017,31(2):142–145.Wang J,Chen F,Wen M X,Li X L,Rao X Y,Xia Z L,Xu B B.Effect of 6-BA treatment on drought tolerance of roasted tobacco.Crop Res,2017,31(2):142–145(in Chinese with English abstract).
[14]耿兵婕,叶苗苗,陈研,王孟昌,马尚宇,黄正来,张文静,樊永惠.外源6-BA和KH2PO4对花后受渍小麦根系抗氧化酶和无氧呼吸酶活性的影响.浙江农业学报,2023,35:2275–2285.Geng B J,Ye M M,Chen Y,Wang M C,Ma S Y,Huang Z L,Zhang W J,Fan Y H.Effects of exogenous 6-BA and KH2PO4 on antioxidant enzymes and anaerobic respiratory enzymes activities in post-flowering impregnated wheat roots.Acta Agric Zhejiangensis,2023,35:2275–2285(in Chinese with English abstract).
[15]刘凯歌,龚繁荣,宋云鹏,张丽丽.外源6-BA对高温胁迫下甜椒幼苗叶绿素荧光参数和抗氧化酶活性的影响.上海农业学报,2020,36(2):19–25.Liu K G,Gong F R,Song Y P,Zhang L L.Effects of exogenous 6-BA on chlorophyll fluorescence parameters and antioxidant enzyme activities of sweet pepper seedlings under high temperature stress.Acta Agric Shanghai,2020,36(2):19–25(in Chinese with English abstract).
[16]杨晓春,朱宗文,张爱冬,吴雪霞,查丁石.外源6-BA对镉胁迫下茄子幼苗生长、光合特性和内源激素含量的影响.上海农业学报,2017,33(5):17–24.Yang X C,Zhu Z W,Zhang A D,Wu X X,Cha D S.Effects of exogenous 6-BA on growth,photosynthetic characteristics and endogenous hormone contents of aubergine seedlings under cadmium stress.Acta Agric Shanghai,2017,33(5):17–24(in Chinese with English abstract).
[17]邹琦.植物生理学实验指导.北京:中国农业出版社,2000.Zou Q.Experimental Guide to Plant Physiology.Beijing:China Agriculture Press,2000(in Chinese with English abstract).
[18]Zou J N,Jin X J,Zhang Y X,Ren C Y,Zhang M C,Wang,M X.Effects of melatonin on photosynthesis and soybean seed growth during grain filling under drought stress.Photosynthetica,2019,57:512–520.
[19]杨媛.不同时期干旱对苎麻生理生化及产量品质的影响.华中农业大学硕士学位论文,湖北武汉,2019.Yang Y.Effects of Different Periods of Drought on the Physiology,Biochemistry and Yield Quality of Ramie.MS Thesis of Huazhong Agricultural University,Wuhan,Hubei,China,2019(in Chinese with English abstract).
[20]李林宇.外源6-BA和褪黑素对干旱胁迫下越橘生理特性影响的研究.吉林农业大学硕士学位论文,吉林长春,2023.Li L Y.Effects of Exogenous 6-BA and Melatonin on Physiological Characteristics of Lingonberry under Drought Stress.MS Thesis of Jilin Agricultural University,Changchun,Jilin,China,2023(in Chinese with English abstract).
[21]王金强,李思平,刘庆,李欢.喷施生长调节剂缓解甘薯干旱胁迫的机理.中国农业科学,2020,53:500–512.Wang J Q,Li S P,Liu Q,Li H.Mechanism of spraying growth regulators to alleviate drought stress in sweetpotato.Sci Agric Sin,2020,53:500–512(in Chinese with English abstract).
[22]王洋,于森淼,王旌扬,宋海龙,雷耀东,张海燕,刘波.干旱胁迫和复水对萱草的生理特性的影响.中国农学通报,2023,39(19):58–64.Wang Y,Yu S M,Wang J Y,Song H L,Lei Y D,Zhang H Y,Liu B.Effects of drought stress and rehydration on physiological characteristics of daylily.Chin Agric Sci Bull,2023,39(19):58–64(in Chinese with English abstract).
[23]刘文瑜,杨发荣,黄杰,魏玉明,李健荣.干旱胁迫对藜麦幼苗生长和叶绿素荧光特性的影响.干旱地区农业研究,2019,37(4):172–177.Liu W Y,Yang F R,Huang J,Wei Y M,Li J R.Effects of drought stress on growth and chlorophyll fluorescence characteristics of quinoa seedlings.Agric Res Arid Areas,2019,37(4):172–177(in Chinese with English abstract).
[24]Yin D M,Chen S M,Chen F D,Guan Z Y,Fang W M.Morphological and physiological responses of two chrysanthemum cultivars differing in their tolerance to waterlogging.Environ Exp Bot,2009,67:87–93.
[25]Biankas S.The role of ethylene and ROS in salinity,heavy metal,and flooding responses in rice.Front Plant Sci,2014,5:685.
[26]Wang N,Chen H,Wang L.Physiological acclimation of Dicranostigma henanensis to soil drought stress and rewatering.Acta Soc Bot Polon,2021,90:907.
[27]胡哲森,许长钦,傅瑞树.锥栗幼苗对水分胁迫的生理响应及6-BA的作用.福建林学院学报,2000,(3):199–202.Hu Z S,Xu C Q,Fu R S.Physiological response of chestnut seedlings to water stress and the role of 6-BA.J Forest Environ,2000,(3):199–202(in Chinese with English abstract).
[28]王明月,陈明,唐雪东,刘炳含,周思佳,陈国双.外源一氧化氮对中度干旱胁迫下越橘幼苗生理生化特性的影响.山东农业科学,2023,55(7):49–56.Wang M Y,Chen M,Tang X D Liu B H,Zhou S J,Chen G S.Effects of exogenous nitric oxide on physiological and biochemical characteristics of lingonberry seedlings under moderate drought stress.Shandong Agric Sci,2023,55(7):49–56(in Chinese with English abstract).
[29]郑桂萍,李金峰,钱永德,吕艳东,刘丽华,王伯伦.农作物综合抗旱性指标的评价分析.中国农学通报,2005,21(10):109–121.Zheng G P,Li J F,Qian Y D,Lu Y D,Liu L H,Wang B L.Evaluation and analysis of comprehensive drought resistance indexes of crops.Chin Agric Sci Bull,2005,21(10):109–121(in Chinese with English abstract).
[30]徐封丰.外源6-苄基嘌呤、水杨酸对高温干旱复合胁迫下半夏植株抗逆性的影响.西南大学硕士学位论文,重庆,2016.Xu F F.Effects of Exogenous 6-benzylpurine and Salicylic Acid on the Resistance of Hemlockia Plants under High Temperature and Drought Compound Stress.MS Thesis of Southwest University,Chongqing,China,2016(in Chinese with English abstract).
[31]Zhang S H,Xu X F,Sun Y M,Zhang J L,Li C Z.Influence of drought hardening on the resistance physiology of potato seedlings under drought stress.J Integr Agric,2018,17:336–347.
[32]Li W T,Ning P,Wang F,Cheng X M,Huang X X.Effects of exogenous abscisic acid(ABA)on growth and physiological characteristics of Machilus yunnanensis seedlings under drought stress.J Appl Ecol,2020,31:1543–1550.
[33]温琦,赵文博,张幽静,梁塔娜,张艳欣,李丽丽,黄凤兰.植物干旱胁迫响应的研究进展.江苏农业科学,2020,48(12):11–15.Wen Q,Zhao W B,Zhang Y J,Liang T N,Zhang Y X,Li L L,Huang F L.Research progress on plant drought stress response.Jiangsu Agric Sci,2020,48(12):11–15(in Chinese with English abstract).
[34]单长卷,郝文芳,张慧成.土壤干早对冬小麦幼苗生理特性的影响.河北农业大学学报,2006,29(4):6–10.San C C,Hao W F,Zhang H C.Effects of soil dryness and early age on the physiological characteristics of winter wheat seedlings.J Hebei Agric Univ,2006,29(4):6–10(in Chinese with English abstract).
[35]李丽杰,顾万荣,孟瑶,王悦力,穆君怡,李晶,魏湜.干旱胁迫下亚精胺对玉米幼苗抗旱性影响的生理生化机制.应用生态学报,2018,29:554–564.Li L J,Gu W R,Meng Y,Wang Y L,Mu J Y,Li J,Wei T.Physiological and biochemical mechanisms of the effect of spermidine on drought tolerance of maize seedlings under drought stress.Chin J Appl Ecol,2018,29:554–564(in Chinese with English abstract).
[36]赵九洲,汤庚国,童丽丽.水分亏缺下SA和6-BA对大花蕙兰的生理调控效应.南京林业大学学报(自然科学版),2004,28(3):27–30.Zhao J Z,Tang G G,Tong L L.Physiological regulation effects of SA and 6-BA on Cymbidium macrorrhizum under water deficit.J Nanjing For Univ(Nat Sci Edn),2004,28(3):27–30(in Chinese with English abstract).
[37]梁颖,胡雪琴,王三根,张宪陶.6-BA对水分胁迫下水稻幼苗生理特性的影响.耕作与栽培,2002,(4):28–29.Liang Y,Hu X Q,Wang S G,Zhang X T.Effect of 6-BA on the physiological characteristics of rice seedlings under water stress.Tillage Cult,2002,(4):28–29(in Chinese with English abstract).
文章摘自:管圣,廖澳,王立琦,等.6-BA调控增强苎麻抗旱性的生理机制研究[J/OL].作物学报,1-13[2024-12-09].
