摘 要:本发明提供了一种亚麻LuNAC73基因及其应用。所述亚麻LuNAC73基因的核苷酸序列如SEQ ID NO.1所示,其编码蛋白的氨基酸序列如SEQ ID NO.2所示。本发明采用病毒诱导的基因沉默技术,通过农杆菌介导法获得LuNAC73基因沉默亚麻株系TRV1:TRV2?LuNAC73(SE)。在SE中,纤维素、半纤维素及木质素合成均受到抑制。LuNAC73基因对亚麻纤维素、半纤维素及木质素的合成均有调控作用,是亚麻纤维发育的重要调控基因,在纤维侵入性伸长和次生壁增厚过程中均发挥重要的调控作用,该基因的发现为改良亚麻纤维产量和品质的分子育种提供了重要基因资源。
权利要求
1.一种调控亚麻纤维发育的LuNAC73基因,其特征在于:所述LuNAC73基因的核苷酸序列如SEQ ID NO.1所示。
2.如权利要求1所述的LuNAC73基因在调控亚麻纤维发育中的应用。
3.根据权利要求2所述的应用,其特征在于:所述调控亚麻纤维发育包括如下一种或多种:
1)通过将亚麻中的LuNAC73基因沉默来降低亚麻纤维素、半纤维素及木质素含量;
2)通过将亚麻中的LuNAC73基因沉默来改变亚麻纤维侵入性伸长生长相关基因表达水平;
3)通过将亚麻中的LuNAC73基因沉默来改变亚麻次生壁合成相关基因表达水平。
4.根据权利要求3所述的应用,其特征在于:所述将亚麻中的LuNAC73基因沉默的方法为:在亚麻LuNAC73基因的序列中选取一段靶序列,利用上游引物LuNAC73F(XbaI)和下游引物LuNAC73R(BamHI)克隆靶序列,通过酶切法将靶序列与pTRV2载体连接,构建VIGS重组载体,将VIGS重组载体导入亚麻植株,从而将亚麻中的LuNAC73基因沉默;
其中,所述靶序列如SEQ ID NO.5所示;
所述上游引物LuNAC73F(XbaI)和下游引物LuNAC73R(BamHI)序列为:
LuNAC73F(XbaI):5’-TCTAGAGAGAAGACCAATTGGGTG-3’,
LuNAC73R(BamHI):5’-GGATCCCTTCAGTTCCTTTGATGAT-3’。
5.如权利要求1所述的LuNAC73基因在亚麻分子育种中的应用。
6.一种调控亚麻纤维发育的方法,其特征在于:所述方法包括将亚麻中的LuNAC73基因沉默,所述LuNAC73基因的核苷酸序列如SEQ ID NO.1所示。
7.根据权利要求6所述的方法,其特征在于:所述调控亚麻纤维发育包括如下一种或多种:
1)降低亚麻纤维素、半纤维素及木质素含量;
2)改变亚麻纤维侵入性伸长生长相关基因表达水平;
3)改变亚麻次生壁合成相关基因表达水平。
说明书
亚麻LuNAC73基因及其在调控亚麻纤维发育中的应用
技术领域
本发明涉及基因工程领域,具体涉及亚麻LuNAC73基因及其应用。
背景技术
在高等植物发育中,细胞体积的增加可以通过三种主要方式进行;协同生长、突出生长和侵入生长。协同生长是最常见的类型,通常表现在伸长区。突出生长只发生在表皮层,并导致毛状体的形成,如根毛或棉花种子纤维的形成。第三种类型是侵入性生长,纤维形成属于此类型。纤维束在植物器官内的发育主要是单个纤维的侵入性伸长生长。在侵入性伸长生长过程中,纤维必须完成的关键过程包括:1.通过合成和分泌细胞壁聚合物和松弛细胞壁扩大细胞壁;2.通过吸水、渗透物积累及膜成分合成增加液泡体积。纤维侵入性伸长生长对植物结构的形成具有重要意义。亚麻是我国重要的纤维作物。亚麻初生韧皮部纤维起源于原形成层,靠近顶端分生组织,在发育的最初阶段,它们与周围组织协同生长。侵入性伸长生长开始于距茎尖1-2mm处,并在茎下持续数厘米,直到“snappoint”点,此位点向下纤维向细胞壁增厚转变。纤维细胞伸长影响纤维产量,次生壁增厚影响纤维品质,鉴定和利用纤维伸长、次生壁增厚相关基因对于解析亚麻纤维发育的分子机理以及提高纤维产量、品质具有重要的理论和实践意义。
本发明采用的技术方案如下:
一种调控亚麻纤维发育的LuNAC73基因,所述LuNAC73基因的核苷酸序列如SEQ IDNO.1所示。
上述一种LuNAC73基因在调控亚麻纤维发育中的应用,所述调控亚麻纤维发育包括如下一种或多种:
1)通过将亚麻中的LuNAC73基因沉默来降低亚麻纤维素、半纤维素及木质素含量;
2)通过将亚麻中的LuNAC73基因沉默来改变亚麻纤维侵入性伸长生长相关基因表达水平;3)通过将亚麻中的LuNAC73基因沉默来改变亚麻次生壁合成相关基因表达水平。
上述将亚麻中的LuNAC73基因沉默的方法为:在亚麻LuNAC73基因的序列中选取一段靶序列,利用上游引物LuNAC73F(XbaI)和下游引物LuNAC73R(BamHI)克隆靶序列,通过酶切法将靶序列与pTRV2载体连接,构建VIGS重组载体,将VIGS重组载体导入亚麻植株,从而将亚麻中的LuNAC73基因沉默;
其中,所述靶序列如SEQ ID NO.5所示;
上游引物LuNAC73F(XbaI)和下游引物LuNAC73R(BamHI)序列为:
LuNAC73F(XbaI):5’-TCTAGAGAGAAGACCAATTGGGTG-3’,
LuNAC73R(BamHI):5’-GGATCCCTTCAGTTCCTTTGATGAT-3’。
上述一种LuNAC73基因在亚麻分子育种中的应用。
一种调控亚麻纤维发育的方法,所述方法包括将亚麻中的LuNAC73基因沉默,所述LuNAC73基因的核苷酸序列如SEQ ID NO.1所示;所述调控亚麻纤维发育包括如下一种或多种:
1)降低亚麻纤维素、半纤维素及木质素含量;
2)改变亚麻纤维侵入性伸长生长相关基因表达水平;
3)改变亚麻次生壁合成相关基因表达水平。
本发明的显著优点在于:
本研究利用病毒诱导的基因沉默(virus-induced gene silencing,VIGS)技术,通过农杆菌介导法获得LuNAC73基因沉默的亚麻株系TRV1:TRV2-LuNAC73(SE)。在SE中,纤维素、半纤维素及木质素合成均受到抑制,与TRV1:TRV2对照植株(CK)相比纤维素含量降低了5.78%(p<0.05),半纤维素含量降低了5.76%(p<0.05),木质素含量降低了7.49%(p<0.05)。结果表明,LuNAC73基因对纤维素、半纤维素及木质素的合成均有调控作用。通过RNA-Seq分析发现,与CK相比,在SE中LuNAC73基因表达量下降了36.69%,多个与侵入性伸长生长相关基因的表达被显著上调或下调表达(|Log2Foldchange|>1,p.adj<0.05),同时在SE中与次生壁合成相关的纤维素合酶LuCesA4、LuCesA8及LuMyb46、LuMyb83、LuNST1、LuNST3、LuSND1等转录调控因子表达量均显著下调(p<0.05)。上述结果表明:LuNAC73(Lus10013967)是亚麻纤维发育的重要调控基因,在纤维侵入性伸长和次生壁增厚过程中均发挥重要的调控作用,该基因的发现为改良亚麻纤维产量和品质的分子育种提供了重要基因资源。
附图说明
图1:亚麻LuNAC73基因沉默植株表型分析。A,WT(未侵染);B:pTRV1:pTRV2-PDS漂白表型;C:pTRV1:pTRV2;D:pTRV1:pTRV2-NAC73。

图2:亚麻LuNAC73基因沉默植株化学组分分析。
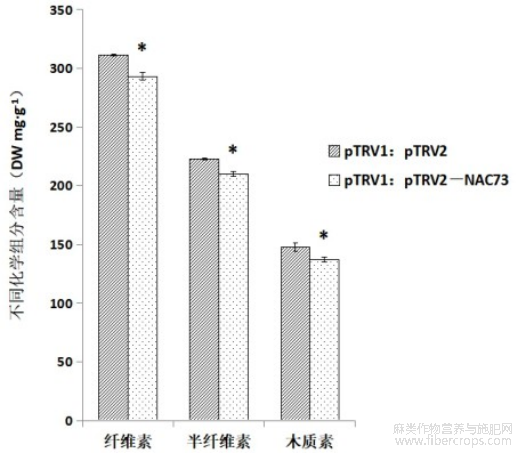
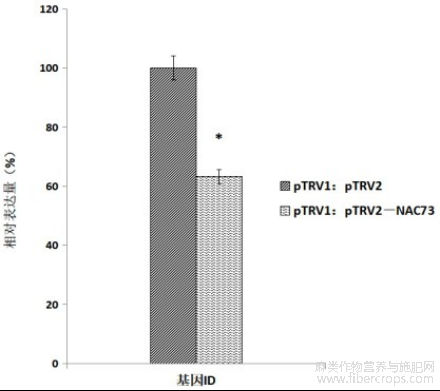
图3:亚麻LuNAC73基因沉默植株基因表达分析。
具体实施方式
下面结合附图和具体实施例对本发明作进一步说明,以使本领域的技术人员可以更好的理解本发明并能予以实施,但所举实施例不作为对本发明的限定。
本发明中所述亚麻LuNAC73(Lu10013967)基因的核苷酸序列如SEQ ID NO.1所示,其编码的蛋白的氨基酸序列如SEQ ID NO.2所示;所述亚麻八氢番茄红素脱氢酶LuPDS(Lus10021967)基因的核苷酸序列如SEQ ID NO.3所示,其编码的蛋白的氨基酸序列如SEQID NO.4所示。
本发明中所用农杆菌为GV3101。
实施例1:
根据Phytozome公共数据库中亚麻LuNAC73(Lu10013967)基因序列设计上下游引物LuNAC73F(XbaI)和LuNAC73R(BamHI),以亚麻cDNA为模板克隆亚麻LuNAC73基因的靶序列(390bp,如SEQ ID NO.5所示),将该靶序列片段插入pTRV2载体的多克隆位点5’-XbaI-BamHI-3’之间,构建VIGS沉默载体pTRV2-NAC73。根据亚麻八氢番茄红素脱氢酶LuPDS(Lus10021967)基因序列设计上下游引物LuPDSF和LuPDSR,以亚麻cDNA为模板克隆亚麻LuPDS基因的靶序列(437bp,如SEQ ID NO.6所示),将该靶序列片段插入pTRV2载体的多克隆位点5’-XbaI-BamHI-3’之间,构建VIGS沉默载体pTRV2-PDS。
将“大紫花”亚麻种子均匀种于营养土中,放置于光照培养箱中,23℃、16h光照,18℃、8h黑暗培养。正常培养10天左右,幼苗长出2片子叶,且子叶展开时进行VIGS试验。VIGS试验方法参照参考文献Chantreau M,Chabbert B,Billiard S,Hawkins S,NeutelingsG.Functional analyses of cellulose synthase genes in flax(Linumusitatissimum)by virus-induced gene silencing[J].Plant Biotechnol J,2015,13(9):1312-1324进行。侵染前提前2天准备菌液。将保存在-80℃农杆菌菌株pTRV1(检测引物pTRV1F/pTRV1R)、pTRV2(检测引物pTRV2F/pTRV2R)、pTRV2-PDS、pTRV2-NAC73菌液PCR检测,检测载体序列正确的菌株在YEB固定培养基(10mg/LKan+50mg/LRif)28℃划线培养2d,然后挑单菌落加1ml YEB液体培养基28℃、200rpm过夜培养,将菌液加入25ml无抗生素YEB液体培养基中28℃、200rpm扩大培养16h,将菌液收集于50ml离心管中,7000rpm离心5min,弃上清液,用新鲜YEB液体培养基(10mM MgCl2+10mM MES+150uM AS)重悬菌体使OD值达到0.6~1.2之间。菌液室温静置3h或者28℃、200rpm振荡培养1h;然后将pTRV1菌液分别与pTRV2菌液、pTRV2-PDS菌液、pTRV2-NAC73菌液等体积混合;选择子叶背面用1ml注射器针头扎三个针眼,注意不要扎透子叶,然后用去掉针头的注射器向叶片背面的针眼里推菌液,注意不要过于用力,菌液不易过多,否则苗后期会死亡。侵染后黑暗培养24h,然后进行正常光照培养(光照16h,黑暗8h)。
表1引物序列

选择SEQ ID NO.5所示的序列作为靶标序列,采用VIGS技术,通过农杆菌介导法获得LuNAC73(Lu10013967)基因沉默亚麻植株。植株的表型如图1所示,侵染的植株与未侵染的植株相比叶片较多。以pTRV1:pTRV2-PDS侵染植株作为指示植株,白化现象表明重组质粒在植物体内表达,茎部白化表型主要出现在子叶以上的第一对真叶与第二对真叶之间;pTRV1:pTRV2对照与pTRV1:pTRV2-NAC73沉默植株(SE)相比表型上无明显差异。
实施例2:
pTRV1:pTRV2、pTRV1:pTRV2-NAC73沉默植株分别取30株,每10株混合做为1次生物学重复,每个样品3次生物学重复。选择长势一致的植株,取材部位为子叶以上第1对与第2对真叶之间的茎段。采用ELISA含量检测试剂盒(齐一,上海)检测样品中的纤维素、半纤维素及木质素含量。
细胞壁化学组分检测结果如图2所示,在LuNAC73基因沉默植株SE中纤维素、半纤维素及木质素的合成均受到抑制。在SE中,纤维素含量为293.2mg/g(DW),与对照相比降低了5.78%(p<0.05);半纤维素含量为209.85mg/g(DW),与对照相比降低了5.76%(p<0.05);木质素含量为136.89mg/g(DW),与对照相比降低了7.49%(p<0.05)。结果表明,LuNAC73基因对纤维素、半纤维素及木质素的合成都有调控作用。
实施例3:
pTRV1:pTRV2、pTRV1:pTRV2-NAC73沉默的植株分别取3株,做为3次生物学重复。选择长势一致的植株取材,取材部位为子叶以上第1对与第2对真叶之间的茎段。将植物样本放入研钵中,加入液氮,迅速研磨成粉末。采用CTAB法提取植物总RNA,使用DNase I去除基因组DNA污染。通过1.0%琼脂糖凝胶电泳检测RNA质量。转录组测序分析由华大基因科技有限公司完成。
利用RNA-Seq技术检测pTRV1:pTRV2对照植株(CK)与pTRV1:pTRV2-NAC73沉默植株(SE)的差异表达基因。在SE沉默株中,靶基因LuNAC73基因(lus10013967)的表达量与对照相比下降了36.69%(p<0.05)(图3),多个与侵入性伸长生长相关基因的表达被显著上调或下调表达(|Log2Foldchange|>1,p.adj<0.05),这些基因涉及激素、信号转导、细胞壁修饰、次生代谢、转录调控等(表2),同时,参与次生壁合成的纤维素合酶LuCesA4、LuCesA8、LuMyb46、LuMyb83、LuNST1、LuNST3、LuSND1等基因在SE中也被显著下调表达(表3)。
表2LuNAC73(lus10013967)调控亚麻纤维侵入性伸长生长相关基因
表3LuNAC73(lus10013967)调控亚麻次生壁合成相关基因
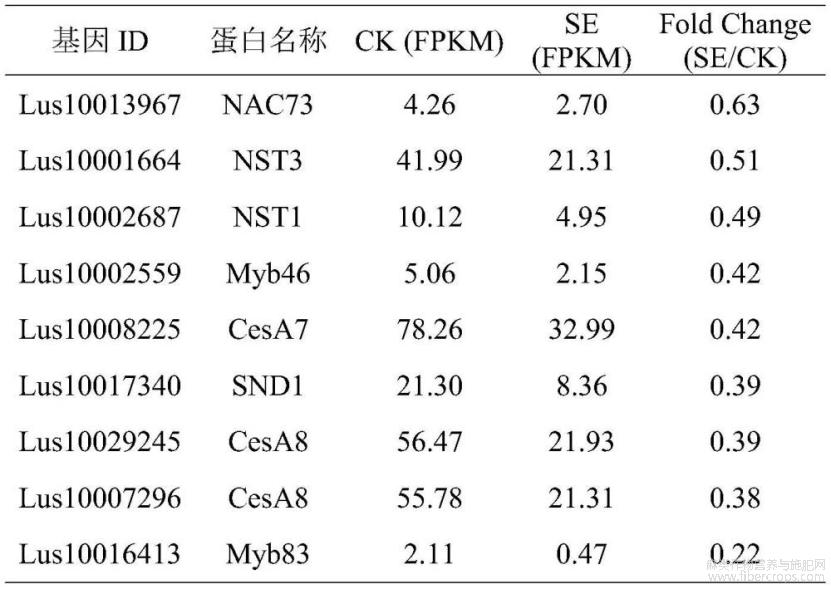
综上,本研究利用病毒诱导基因沉默(VIGS)的方法证明LuNAC73(lus10013967)基因是亚麻纤维发育中的重要调控基因,能够调控纤维素、半纤维素及木质素的合成。转录组差异基因表达分析发现,该基因参与纤维侵入性伸长生长的调控,同时也参与纤维次生壁合成的调控。该基因的发现为提高亚麻纤维产量和改良亚麻纤维品质的分子育种提供了重要的基因资源。
以上所述仅为本发明的较佳实施例,凡依本发明申请专利范围所做的均等变化与修饰,皆应属本发明的涵盖范围。
摘自国家发明专利,发明人:袁红梅 郭文栋 刘丹丹 唐立郦 姚玉波 程莉莉 杨洌 吴广文 康庆华 宋喜霞,申请号:CN202311771569.3,申请日:2023.12.21
