摘 要:粒重性状对于胡麻育种具有重要意义。本研究以胡麻小粒型材料CLX为母本,大粒型材料BLX为父本,杂交获得F1和F2群体,应用主基因–多基因遗传模型对胡麻籽粒千粒重、籽粒表面积、籽粒周长、籽粒长度和籽粒宽度的遗传效应进行了分析。结果表明,粒重的遗传模型为2对加性主基因+加性–显性多基因控制,主基因遗传率为84.59%,多基因遗传率为14.73%。籽粒表面积、籽粒长度、籽粒宽度受2对加性–显性–上位性主基因+加性–显性多基因控制,3个性状的主基因遗传率分别为45.69%、37.09%、19.27%。籽粒周长受2对等显性主基因+加性–显性多基因控制,主基因遗传率为68.24%。胡麻粒型性状的遗传以主基因遗传效应为主,可通过杂交在后代群体中早代选择进行改良利用。
关键词:胡麻;籽粒性状;遗传模型
粒重是作物产量构成的重要因素之一,较高的千粒重是实现作物高产和品质的保证,而籽粒面积、周长、粒长、粒宽等表型性状是影响粒重的主要构成因素[1]。前期研究[2,3]表明,种子大小会对作物的产量和品质产生影响,千粒重越高的胡麻籽粒中含油率和亚麻酸含量越高;干瘪短小的胡麻籽粒中亚油酸和木酚素含量越高。粒重越大越饱满的种子,种子活力和田间出苗能力越强,苗期植株更健壮,产生的分蘖更多,从而形成更多的果穗数,为高产打下基础[4,5]。因此,粒重和粒型性状的深入研究对于提高胡麻产量具有重要意义。
籽粒大小、形态等籽粒特性遗传基础复杂,是典型的数量性状,受多基因控制,且以加性效应为主,在分离群体中表现为连续变异,变异频率常呈整体分布[6]。盖钧镒、章元明等提出的主基因+多基因混合遗传模型在油菜、胡麻等油料作物数量性状上研究应用比较多[7,8,9]。但是,关于胡麻中籽粒相关性状基因的分布与遗传分析却鲜见报道。本文运用主基因+多基因混合遗传模型分析了2个亲本群体、F1和F2群体的千粒重、籽粒表面积、籽粒周长、粒长、粒宽等性状的遗传机制,旨在为改良胡麻籽粒重,提高产量奠定基础。
1 材料与方法
1.1 试验材料
以CLX(小粒型材料)为母本,BLX(大粒型材料)为父本,杂交获得F1代,F1自交得到F2代单株群体,试验材料由甘肃省农业科学院作物所提供。
1.2 试验方法
2022年3月上旬将上述P1、P2、F1和F2群体种植在甘肃省农业科学院兰州试验地。行长2m,每行播种300粒,行距20cm。亲本P1、P2,F1每个材料种植5行,F2种植10行。整个生育期常规管理。待实验材料成熟后单株收获脱粒,晾干后用万深SC-G自动考种分析及千粒重分析系统测量籽粒千粒重(1000-seed weight,TSW)、籽粒表面积(Seed surface area,SSA)、籽粒周长(Seed circumference,SC)、籽粒长度(Seed length,SL)、籽粒宽度(Seed width,SW)。
1.3 数据处理
利用主基因+多基因遗传模型对胡麻籽粒千粒重、表面积、周长、长度和宽度进行遗传分析[10,11]。通过SEA-G4F2算法计算模型参数,判断AIC值并通过适合性检验,选择每个性状的遗传模型,并估算各性状的遗传参数。
2 结果与分析
2.1 胡麻F2群体粒型性状分析
F15个性状的平均值介于两个亲本之间,但更接近于BLX各性状的值;F2群体千粒重的变异系数最大(23.35%),其次是籽粒表面积(17.49%),籽粒周长、籽粒长度、籽粒宽度的变异系数分别为9.00%、8.82%、9.49%(表1)。说明分离世代群体籽粒性状表型变异丰富,明显高于P1、P2和F1群体,且有超亲现象。籽粒千粒重、表面积、周长、长度和宽度性状均呈连续性分布,为右偏离正态分布(图1)。
表1 F2群体粒型性状的统计值
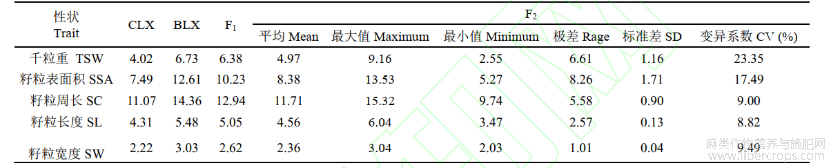
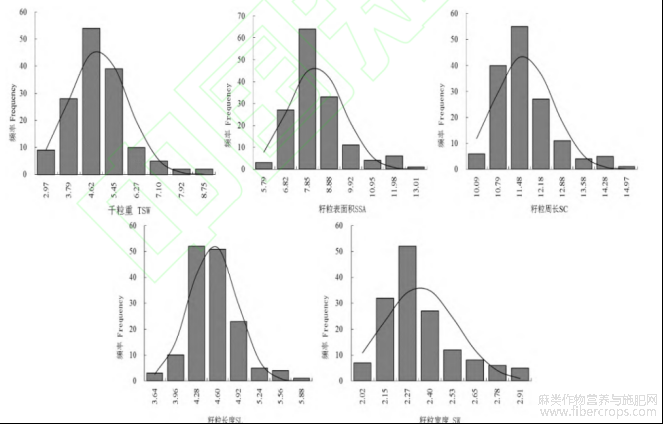
图1 F2群体不同粒型性状的分布图
2.2 不同籽粒性性状遗传模型选择及适合性检验
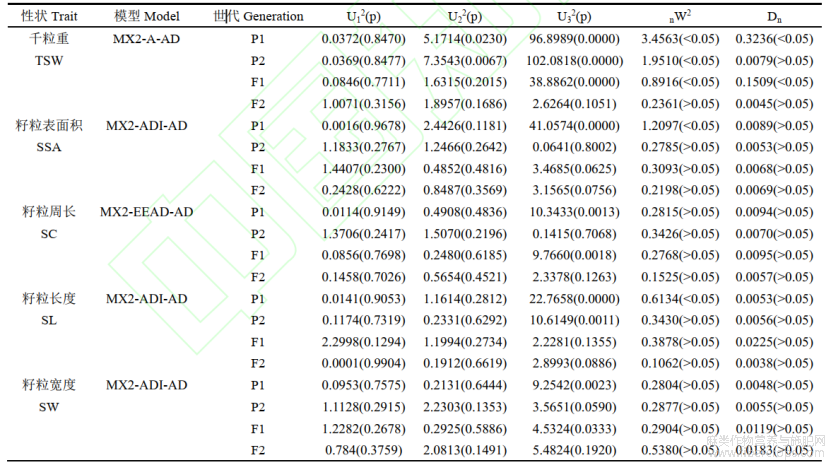
运行SEA-G4F2软件,对5个籽粒性状的表型数据进行模型分析,首先得到不同粒型性状模型的极大似然函数值(MLV)和AIC值(表2)。根据AIC值最小原则从中选择3个模型作为备选模型。对3个模型的4个群体分别进行适合性检验,U12、U22、U32、Smirnov检验(nW2)和Kolmogorov检验(Dn)(表3),每个性状的最优模型为3个备选模型中统计量达到显著水平个数最少的模型。粒重的最适遗传模型为MX2-A-AD,受2对加性主基因+加性–显性多基因控制;籽粒表面积、籽粒长度、籽粒宽度的最适遗传模型为MX2-ADI-AD,受2对加性–显性–上位性主基因+加性–显性多基因控制;籽粒周长的遗传模型为MX2-EEAD-AD,受2对等显性主基因+加性–显性多基因控制。
表2 不同遗传模型下5个籽粒性状的极大似然函数值和AIC值
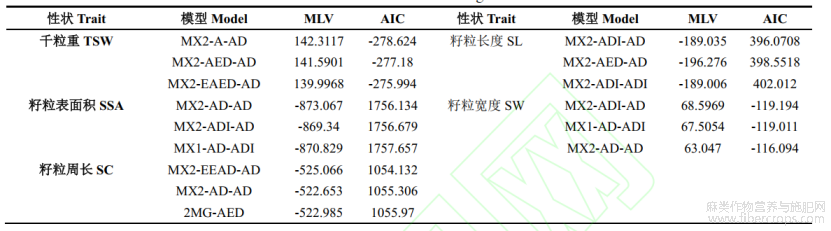
表3 5个籽粒性状遗传模型的适合性检验
2.3 遗传参数估计
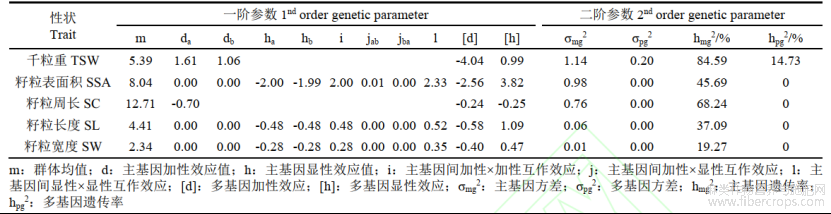
采用最小二乘法估算粒重和4个粒型性状遗传模型的一阶、二阶遗传参数(表4),5个性状均受主基因+多基因遗传控制。籽粒千粒重受两对加性主基因+加性–显性多基因控制,2对粒重主基因都具有正向加性效应,多基因的加性效应值为负向,显性效应值为正向;千粒重主基因遗传率为84.59%,多基因遗传率为14.73%。籽粒表面积、籽粒长度、籽粒宽度受2对加性–显性–上位性主基因+加性–显性多基因控制,主基因的显性效应值为负向,两对主基因之间的加性×加性互作效应值和显性×显性互作效应值均为正向;多基因加性效应值为正向,多基因显性效应值为负向;3个性状的主基因遗传率分别为45.69%、37.09%、19.27%。籽粒周长受2对等显性主基因+加性–显性多基因控制,主基因加性效应值为负向,多基因加性效应值和显性效应值均为负向;主基因遗传率为68.24%。
表4 5个胡麻籽粒性状的遗传参数估计值
3 讨论
粒重和粒型是影响作物产量和品质的重要因素,是由多基因控制的复杂数量性状[12]。影响粒重的因素主要包括籽粒面积、周长、粒宽、粒长等粒型因素[13]。粒重和粒型性状是高产、优质胡麻品种选育的重要性状和改良性状,深入解析胡麻粒重和粒型性状的遗传机理是选育高产优质胡麻品种的基础。本研究表明,粒重受2对加性主基因+加性–显性多基因控制,2对主基因的加性效应为正向,多基因的加性效应值为负向,显性效应值为正向;主基因遗传率高达84.59%,是5个性状中主基因遗传力最高的。但本研究结果与袁谦在小麦中粒重受两对加性–显性–上位性主基因+加性–显性–上位性多基因遗传模型稍有不同[14]。胡麻千粒重的改良应该在早期进行选择,利用主基因加性效应的不断累积来选育大粒品种。水稻粒长和粒宽受主基因和微效基因共同调控,其遗传力较高,一般认为水稻粒型主要是由多基因共同控制[15]。胡麻籽粒表面积、粒长、粒宽受2对加性–显性–上位性主基因+加性–显性多基因控制,与水稻研究结果基本一致;主基因的显性效应为负值,两对主基因之间的互作效应值均为正向;多基因加性效应值为正向,显性效应值为负向;3个性状的主基因遗传率分别为45.69%、37.09%、19.27%。胡麻籽粒表面积和粒长的遗传模型与小麦和玉米上研究结果不同[16],而粒宽遗传模型与小麦遗传模式相同[14]。籽粒周长受2对等显性主基因+加性–显性多基因控制,主基因加性效应值、多基因加性效应值和显性效应值均为负向;主基因遗传率为68.24%,主基因遗传力仅次于粒重,但与小麦籽粒周长为两对加性–显性–上位性主基因遗传模型结果不同[14]。可见,粒重和粒型的遗传模型受不同作物、不同群体和不同环境的影响比较大。本研究中粒重、籽粒周长、粒表面积、粒长的主基因遗传力均比较高,表明这些性状遗传比较稳定,受环境条件的影响小,可利用杂交在后代群体中早代选择进行改良。本研究应用主基因+多基因模型分析方法,研究胡麻粒重及4个粒型性状的遗传规律,为胡麻新品种选育中籽粒性状的改良提供参考。
4 结论
胡麻粒重和4个粒型性状呈正态分布,通过主基因+多基因遗传模型分析表明,千粒重受2对加性主基因+加性–显性多基因控制,2对主基因的加性效应为正向,起正向调控作用;多基因的加性效应值为负向,显性效应值为正向;籽粒表面积、籽粒长度、籽粒宽度受两对加性–显性–上位性主基因+加性–显性多基因控制,主基因的显性效应为负值,两对主基因之间的互作效应值均为正向;多基因加性效应值为正向,显性效应值为负向;籽粒周长受2对等显性主基因+加性–显性多基因控制,主基因加性效应值、多基因加性效应值和显性效应值均为负向。粒重、籽粒表面积、籽粒周长、籽粒长度4个性状的主基因遗传率比较高,基因间互作效应值有正有负,遗传基础比较复杂。
参考文献
[1]王瑞霞,张秀英,伍玲,等.不同生态环境下冬小麦籽粒大小相关性状的QTL分析.中国农业科学,2009,42(2):398-407.
[2]赵利,王斌,苗红梅,等.胡麻种质资源籽粒表型与品质性状评价及其相关性研究.植物遗传资源学报,2020,21(1):243-251.
[3]刘栋,马琴,李爱荣,等.亚麻种质资源种子形态性状与含油量的分析与评价.作物杂志,2020(3):34-41.
[4]吉春容,李世清,李生秀.品种、种子大小和施肥对冬小麦生物学特性的影响.生态学报,2007,27(6):2498-2506.
[5]刘生祥,宋晓华.春小麦种子大小对主要性状及产量的影响.种子,2003(1):26-27.
[6]陈冰蠕,石英尧,崔金腾,等.利用BC2 F 2高代回交群体定位水稻籽粒大小和形状QTL.作物学报,2008,34(8):1299-1307.
[7]王利民,党照,赵玮,等.胡麻亚麻酸含量的遗传分析.西北农业学报,2020,29(6):942-948.
[8]王斌,王利民,张建平,等.胡麻RIL群体苗期抗旱性状的主基因+多基因遗传分析.干旱地区农业研究,2018,36(5):14-20.
[9]刘霞,张冰冰,马兵,等.甘蓝型油菜株高及其相关性状的主基因+多基因遗传分析.西北农业学报,2018,27(4):528-536.
[10]盖钧镒.植物数量性状遗传体系的分离分析方法研究.遗传,2005,27(1):130-136.
[11]章元明.植物数量遗传学的建立、发展与展望.南京农业大学学报,2012,35(5):19-24.
[12]Wang G,Wang Y,Ying J,et al.Identification of qLG2, qLG8, and qWG2 as novel quantitative trait loci for grain shape and the allelic analysis in cultivated rice.Planta,2020,252(2):316.
[13]Kang Y,Zhang M,Zhang Y,et al. Genetic mapping of grain shape associated QTL utilizing recombinant inbred sister lines in high yielding rice (Oryza sativa L.).Agronomy,2021,11(4):705.
[14]袁谦,赵永涛,张中州,等.小麦籽粒性状的遗传效应分析及其育种策略. 麦类作物学报,2023,43(4):434-441.
[15]丁丹.水稻5个粒型相关基因的分子标记开发与效应分析.南京:南京农业大学,2014.
[16]张中伟,杨海龙,付俊,等.玉米粒长性状主基因+多基因遗传分析.作物杂志,2019(5):37-40.
文章摘自:王斌,赵利,侯静静,刘杰,段艳巧.胡麻粒重和粒型性状的主基因-多基因遗传模型分析[J/OL].作物杂志.https://link.cnki.net/urlid/11.1808.S.20231208.1450.004
