摘 要: 为探究水分胁迫下外源氯化钙(CaCl2)对亚麻幼苗生长及生理的影响,采用盆栽控水法,以“双亚12”幼苗为试验材料,设置0(CK)、1(Ca1)、3(Ca3)、6(Ca6)、12(Ca12)和24mmol/L(Ca24)等6个CaCl2浓度处理,于枞形期根施处理。结果表明,Ca6与Ca12处理能够有效缓解水分胁迫对亚麻幼苗形态的不利影响。与CK相比,在干旱胁迫下,Ca6与Ca12显著增加了株高、茎粗、叶片数、地上部和地下部鲜重,增加范围在27.10%~108.44%之间;在水淹胁迫下,Ca6与Ca12显著增加了株高、根粗、叶片数和地上部、地下部鲜重,增加范围在15.13%~66.29%之间。CaCl2对胁迫结束时的抗性生理酶活性状态表现不同,在干旱胁迫下,Ca6和Ca12比CK显著增加了PAL和CAT活性,显著降低了SOD和POD活性;在水淹胁迫下,Ca6与Ca12比CK显著增加CAT活性,显著降低了SOD、POD和PAL活性。Ca6和Ca12处理在胁迫结束时均显著降低了膜脂过氧化物质,与CK相比,Ca6处理显著降低了干旱胁迫与水淹胁迫下的O??含量和MDA含量,分别为33.93%和56.05%与98.18%和76.17%;Ca12处理显著降低了H2O2含量,分别为24.14%(干旱)和25.16%(水淹)。变异系数分析分析表明,Ca6处理的综合影响最大,分别为20.07(干旱)和25.11(水淹)。隶属函数分析表明,Ca6处理的平均得分最高,分别为0.75(干旱)和0.63(水淹)。综上,6~12mmol/L的CaCl2预处理亚麻幼苗可有效缓解水分胁迫对亚麻幼苗形态和生理的不利影响,其中6mmol/L的CaCl2是最适预处理浓度。同时,本研究也证明了CaCl2缓解干旱和水淹胁迫的最适浓度是同一浓度。
关键词:CaCl2;水分胁迫;亚麻幼苗;抗性生理;形态
亚麻(Linum usitatissimum L.)是我国重要的区域特色经济作物之一,是保护土壤的重要轮作作物之一,也是人们生活所需的主要纺织原料和油料作物之一[1]。其茎秆和籽粒广泛被用于人们的医、食、衣、行。我国亚麻主要种植在西北和东北地区,该地区的生产总量不足全国总需求量的20%。其中,这些地区常常受到极端气候的影响是限制这些地区亚麻生产的重要因素。在亚麻不到80d的生育期内,遭受干旱和水淹(水渍)等自然灾害已成为这些地区亚麻生产上亟待解决的问题[2]。
作物在逆境胁迫下的稳产途径有很多,如选育抗逆新品种、适时播种、补充外源生长调节剂或营养元素等。其中,补充外源营养元素(N?P?K?Ca?B?Cu等)是降低、缓解和消除逆境对作物伤害的快速、高效、低成本的措施。钙(Ca2+)是植物体内重要的营养元素,不仅是信号传导的“第二信使”,还是提升作物适应逆境胁迫能力的关键元素[3]。有研究[4-5]表明,补充适量的外源Ca2+不仅在正常水分下,促进种子萌发和植株生长发育,还在干旱或水淹时有效缓解水分胁迫对作物的伤害。外源Ca2+主要通过提升抗氧化酶系统的酶活性,降低MDA等有害物质的含量,缓解水分胁迫对作物的不利影响[6]。大多数作物对水分胁迫下CaCl2响应的最适浓度是10mmol/L[7-9]。而有的研究表明,不同处理方式,不同基因型的花生对干旱胁迫下CaCl2响应的最适浓度是不同的[10-11],也有研究表明玉米对干旱和水淹胁迫下Ca2+的响应浓度分别是10mm和5.5mm[7,12]。还有研究表明,不同化合态的钙虽然缓解紫花苜蓿水淹胁迫的最适响应浓度5mmol/L,但其缓解的生理机制不同[13]。此外,还有研究表明,对于外源钙施用时间来说,预处理效果优于补施效果[12]。
为此,本研究从重度干旱胁迫和水淹胁迫的两个极端水分胁迫的角度,通过在亚麻枞形期进行不同浓度钙肥的灌根处理,探讨根施钙对亚麻形态、抗性生理酶活性及膜脂过氧化物质等指标的影响,结合变异系数、Spearman相关系数和隶属函数分析方法,明确亚麻在水分胁迫下的CaCl2最适施用浓度,以期为亚麻优质和高效生产提供理论依据及技术参考。
1 材料与方法
1.1 试验材料与方法
将150粒“双亚12”的亚麻种子播种于装有3kg大豆田耕层土的花盆中,内径为25cm,高为18cm,在黑龙江省科学院大庆分院人工气候室内进行培养,温度为18~25℃,光周期为14/10h(昼/夜)。待第5对真叶完全展开时,定苗60株,定苗标准为:长势一致、无病虫害、无机械损伤。待第7对真叶完全展开时(枞形期),进行CaCl2液体预处理,处理量为100mL/盆,此时记为第0天,并于第1天开始进行水分胁迫处理,共胁迫15d。
本试验所设置的水分胁迫是参考邓欣[14]的研究并通过预实验得出:干旱胁迫以重度干旱胁迫(即田间最大持水量的35%~45%)为标准;水淹胁迫以淹没第1对真叶(即水深2cm)为标准。CaCl2预处理的浓度分别为:0?1?3?6?12和24mmol/L,共计6个处理,(以下简称CK?Ca1?Ca3?Ca6?Ca12和Ca24)每个处理3次重复。在水分胁迫第15d时,每盆亚麻幼苗的一半用于测定生理指标,一半备选,用来统计鲜重、株高、茎粗、根粗和叶片数等形态参数。
1.2 形态参数的测定
将亚麻植株清洗干净后,每个处理选取长势一致且完整的15株进行测定形态。
使用统计法测定叶片数,只统计完全展开的成熟叶片,其中发黄未掉落的叶片记为0.5片。
利用直尺测定株高,即子叶痕至植株顶端的距离。
利用数显游标卡尺测定茎粗和根粗,即株高的1/3处为茎粗,下胚轴白色和绿色相交处为根粗。
利用千分之一天平测定地上部和地下部鲜重,总鲜重为地上部和地下部鲜重之和。
1.3 抗性生理酶活性及膜脂过氧化产物的测定
采用NBT比色法测超氧化物歧化酶活性(SOD);采用愈创木酚法测过氧化物酶活性(POD);采用紫外吸收法测过氧化氢酶活性(CAT);采用紫外吸收法测苯丙氨酸解氨酶活性(PAL)[15]。采用硫代巴比妥酸显色法测丙二醛含量(MDA);采用羟胺法测超氧阴离子含量(O??);采用硫酸钛法测过氧化氢含量(H2O2)[16]。
1.4 数据统计的方法
采用Excel进行数据统计与初步分析,利用Granph Prism 8.0软件科研绘图,采用Tukey法进行单因素方差分析,利用Spearman相关系数进行矩阵相关性分析。
隶属函数计算方法参考公式[17]
(1)同一指标的正向隶属函数=(该指标的测定值–该指标最小值)/(该指标最大值–该指标最小值)
(2)同一指标的反向隶属函数=1–(该指标的测定值–该指标最小值)/(该指标最大值–该指标最小值)
变异系数计算方法参考文献[18],变异系数(Cv)=标准差(CaCl2处理观察值与对照观察值)/平均值(CaCl2处理观察值与对照观察值)×100。
2 结果与分析
2.1 不同浓度的CaCl2对水分胁迫后亚麻形态的影响
由图1-A可知,水分胁迫(CK)使亚麻幼苗上部的叶片呈收敛状态,干旱胁迫主要表现在植株顶端,水淹胁迫主要表现在上1/3部。随着CaCl2浓度的增加,其变化呈“先舒展再收敛”的状态,其中,Ca6处理的亚麻幼苗的植株顶端舒展程度最大。此外,在水淹胁迫下,随CaCl2浓度的增加,叶片大小呈“先变大后变小”的趋势,其中,Ca6处理的叶片最大。
由图1-B、C可知,在水分胁迫下,随着CaCl2浓度的增加,亚麻幼苗的鲜重呈“先增加后降低”的趋势,而不同浓度CaCl2对不同部位的增加效果不同。与对照(CK)相比,在干旱胁迫下(图1-B),Ca6处理对地上部和地下部鲜重的增加效果最大,分别显著增了50.52%和108.44%;在水淹胁迫下(图1-C),Ca12处理对地上部鲜重的增加效果最大,显著增加了29.55%,Ca6处理对地下部鲜重的增加效果最大,显著增加了66.29%。
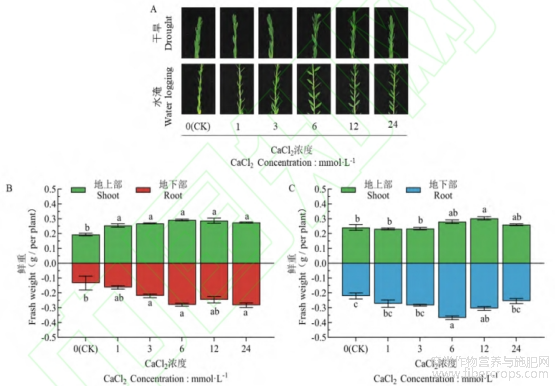
图1 CaCl2对干旱和水淹胁迫后形态的影响
不同小写字母表示处理间存在显著性差异(P<0.05),下同。
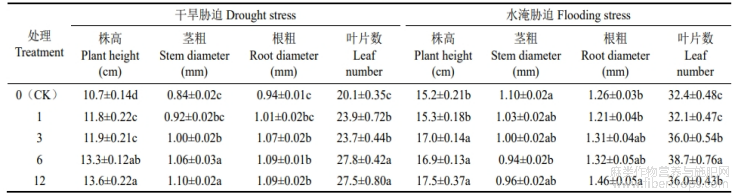
由表1可知,不同浓度CaCl2对水分胁迫下亚麻幼苗的表型参数产生了不同的影响。其中,与对照(CK)相比,在干旱胁迫下,Ca6处理对叶片数影响最大,显著增加了38.31%;Ca12处理对于株高和茎粗影响最大,分别显著增加了27.10%和31.42%;Ca24处理对根粗影响最大,显著增加了31.63%;在水淹胁迫下,Ca6处理对叶片数和茎粗影响最大,显著增加了19.44%的叶片数,而显著降低了14.82%的茎粗;Ca12处理对株高和根粗影响最大,分别显著增加了15.13%和15.87%。
表1 CaCl2对水分胁迫下形态建成指标的影响
![]()
不同小写字母表示处理间存在显著性差异(P<0.05),下同。
2.2 CaCl2对水分胁迫下亚麻幼苗抗性生理酶活性的影响
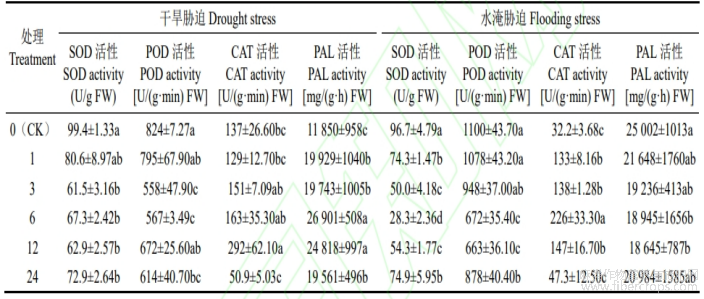
由表2可知,在水分胁迫下,SOD和POD活性随着CaCl2浓度的升高呈“先降低后平缓”的趋势,而CAT则是随着CaCl2浓度的升高呈“先升高后降低”的趋势。其中,与对照相比,在干旱胁迫下,SOD和POD活性在Ca3处理时达到最小,分别显著降低了47.70%和32.28%,CAT活性在Ca12处理时达到最大,显著增加了113.14%;在水淹胁迫下,SOD活性在Ca6处理时值最小,显著降低了70.73%,POD活性在Ca12处理时值最小,显著降低了39.73%,CAT活性在Ca6处理时达到最大,显著升高了601.86%。对于PAL来说,在干旱胁迫下其变化同CAT活性变化规律一致,与CK相比,在Ca6处理时到最大,显著增加了127.01%;而在水淹胁迫下其变化同POD变化规律一致,与CK相比,在Ca12处理时达到最小,显著降低了25.43%。
表2 CaCl2对干旱和水淹胁迫抗性生理酶活性的影响
2.3 CaCl2对水分胁迫下亚麻膜脂过氧化物质的影响
由图2可知,在水分胁迫下,亚麻幼苗叶片的膜脂过氧化的物质随着CaCl2浓度的升高呈“先降低后升高”的趋势。在干旱胁迫下,与对照相比,O??含量(图2-A)和MDA含量(图2-C)在Ca6处理时降低程度最大,分别显著降低了33.93%和56.05%,H2O2含量(图2-B)在Ca12处理时下降的幅度最大,显著下降了24.14%。在水淹胁迫下,与对照相比,O??含量(图2-D)和MDA含量(图2-F)也是在Ca6处理时下降幅度最大,分别显著降低了98.18%和76.17%,H2O2含量(图2-E)也是在Ca12处理时达到最低,显著降低了25.16%。
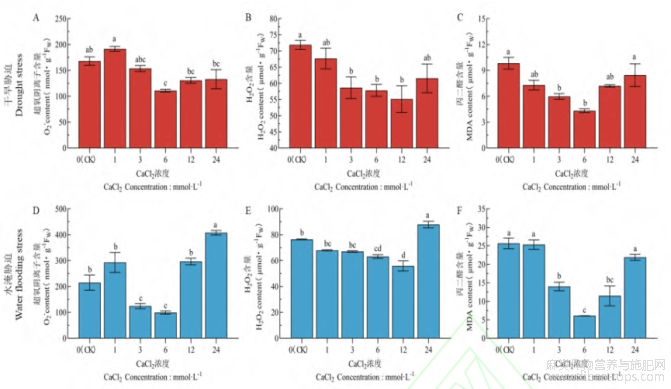
图2 CaCl2对水分胁迫下亚麻膜脂过氧化物质的影响
2.4 变异系数和相关性分析
由图3可知,针对不同氯化钙与亚麻形态和生理指标间进行变异系数分析,结果显示(图3A),两种胁迫下均是Ca6处理平均变异系数最高。其中,在水淹胁迫下,Ca6处理影响最强的指标是总鲜重、地下部鲜重、叶片数、茎粗、SOD活性、CAT活性、MDA含量、O??含量等8个指标;在干旱胁迫下,Ca6处理影响最强的指标是总鲜重、地上部鲜重、叶片数、PAL活性、MDA含量、O??含量等6个指标。针对植株形态与生理各指标间进行Spearman相关性分析,结果显示,在水淹胁迫下,呈强的正相关关系的是:茎粗与SOD活性、POD活性、PAL活性和MDA含量之间,CAT活性与地上部鲜重、总鲜重、地下部鲜重和根粗之间;呈强的负相关关系的是:POD活性、PAL活性和MDA含量与除了茎粗和根粗外其他各个形态指标之间,总鲜重、地下部鲜重和根粗与SOD活性和H2O2含量之间,根粗与PAL活性和MDA含量之间。在干旱胁迫下,株高、地上部鲜重和叶片数与PAL活性呈强的正相关关系;呈强的负相关关系的是:SOD活性与株高之间,POD活性与总鲜重之间,MDA含量与地上部鲜重之间,H2O2含量与除了茎粗和地下部鲜重以外的各个形态指标之间,O??含量与处理茎粗之外的各个形态指标之间。
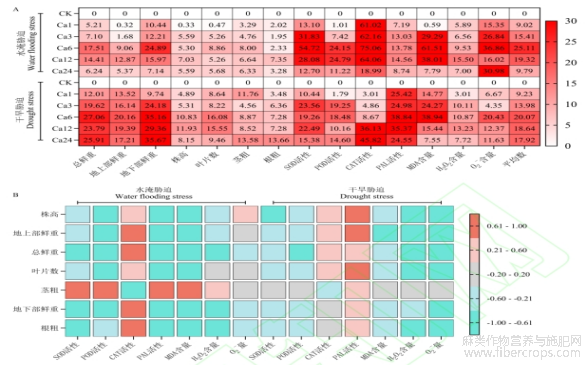
图3 CaCl2对水分胁迫下亚麻幼苗形态指标及生理指标综合分析
图A表示CaCl2浓度与形态指标和生理指标间的变异系数,图B表示形态指标与生理指标间的相关关系。
2.5 综合评价
为综合评价各浓度CaCl2对水分胁迫下亚麻幼苗的缓解效果,通过隶属函数值分析(图4)可知,在水淹胁迫下,Ca6处理的总鲜重、地下部鲜重、叶片数、CAT、MDA、超氧阴离子等6个指标得分为“1”;在干旱胁迫下,Ca6处理的总鲜重、地上部鲜重、叶片数、PAL、MDA、超氧阴离子等6个指标得分为“1”,两种胁迫下均是Ca6处理平均得分最高。
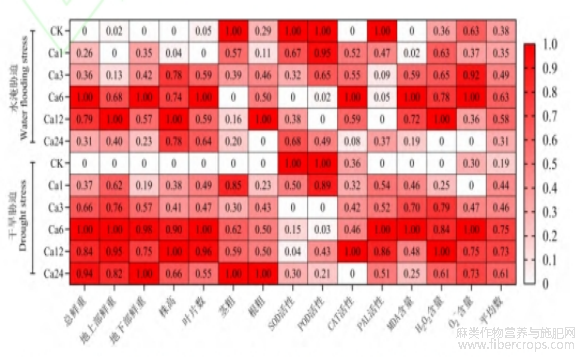
图4
3 讨论
根施适宜浓度或者适宜量的钙(适宜钙)对作物的生长有促进作用。何敏敏等[19]研究表明,适宜钙促进了甜菜地上部和地下部的形态生长发育。么传训等也从根系发育的角度证明了适宜钙是通过促进根系发育,提升了花生的产量。此外,赵海洋等[20]研究表明,适宜钙促进了干旱胁迫下金莲花幼苗叶的生长,范小玉等[21]研究也表明,适宜钙可以促进干旱胁迫下花生的生物量。本研究结果表明,适宜钙促进了干旱和水淹胁迫下亚麻幼苗生物量的积累(图1-B),促进了株高、根粗和叶片数的增加(表1),还使得叶片舒展变大(图1-A)。此外,本研究还发现,在水分胁迫下,适宜钙对地下部生物量的促进效应大于对地上部生物量的促进效应(图1-B),这与前人研究结果相似。说明,根施适宜钙可以促进两种极端水分胁迫下亚麻幼苗生长发育,主要通过促进地下部的生长,缓解水分胁迫对亚麻幼苗生长的不利影响。
抗氧化防御是植物抵御氧化胁迫的一种重要潜在机制,主要由SOD、POD、CAT和APX等酶类组成,由ROS(O???H2O2)和MDA等膜质过氧化物质所体现[7-13]。Zhang等[22]研究表明,CaCl2可有效地提高干旱胁迫下黄花红砂植株的渗透调节物质含量,抗氧化酶活性及相关基因表达,清除过量的ROS,缓解干旱胁迫造成的氧化损伤。Wadood等[23]研究表明,CaCl2是通过进一步提高水分胁迫下大麦SOD、POD和CAT活性,促使大麦体内H2O2含量下降,而Zhu等[24]研究表明,CaCl2是进一步提高胁迫条件下油菜幼苗SOD和APX活性,反而降低了CAT和POD活性,减少水分胁迫对油菜幼苗的伤害的。本研发现,适宜浓度的CaCl2提升了水分胁迫下亚麻幼苗的CAT活性,降低了SOD和POD活性,降低了O??、H2O2和MDA等含量。这可能是每种植物自身的抗氧化酶表达不均衡,致使其在遭受水分胁迫时ROS代谢出现了紊乱,导致ROS积累和MDA积累,由于CAT是亚麻在水分胁迫时清除H2O2的主要限制酶,CaCl2通过激发调节各抗氧化酶均衡发挥作用某个因子(均衡因子),使亚麻体内ROS代谢畅通,进而减少了ROS对亚麻的毒害。至于植物体内的抗氧化酶表达的均衡因子是什么,有待进一步研究。
本研究发现,适宜钙使亚麻在水分胁迫下增加了株高、叶片数和总鲜重(表1),促进叶片舒展,还促进水淹胁迫下亚麻幼苗变大(图1-A)。株高、叶片数和总鲜重与POD呈强的负相关关系,与PAL分别在干旱胁迫和水淹胁迫条件下呈强的正相关关系和强的负相关关系(图3-B),还有水淹胁迫与干旱胁迫CK的PAL测定值的差值大于适宜钙处理下的测定值相差不大(表1)。有研究[25-26]表明,PAL和POD是作物合成木质素、纤维素、和半纤维素等结构糖的关键酶,其活性大小也反映出作物的抗性强弱和形态发育程度。徐荣琼等[27]发现适宜钙处理可促进木质素、纤维素、和半纤维素含量增加,有助于防止植株发生形变。Porakala等[28]研究表明,适宜钙处理促进了营养器官和生殖器官的发育。这可能是干旱胁迫导致POD活性增强和PAL活性降低不能正常产生结构糖,使叶片的支撑力下降从而导致叶片萎蔫和收敛;水淹胁迫导致POD活性增强和PAL活性一样增强,产生过多的结构糖,使细胞无法变大,从而导致叶片变小和收敛。适宜钙通过调节亚麻叶片的这两种酶趋于稳态,从而缓解水分胁迫对亚麻叶片形态的不正常发育。对于适宜钙是否通过促进水分胁迫下亚麻纤维素和木质素发育增强亚麻抗逆性的结果,本试验只是涉及到部分指标,有待于进行其他指标的辅助验证。
变异系数值可直观的表现出胁迫组与正常组之间的影响程度,同样也可以反映处理组与对照组之间的影响程度[7,29-30]。隶属函数可直观地评价处理组与对照组的得分,更加准确地比较处理组之间的得分排名情况[31-33]。本研究发现氯化钙对水分胁迫下亚麻幼苗的影响强弱程度是Ca6>Ca12>Ca24>Ca3>Ca1>CK(干旱),Ca6>Ca12>Ca3>Ca24>Ca1>CK(水淹);平均隶属函数值由大到小的是Ca6>Ca12>Ca24>Ca3>Ca1>CK(干旱),Ca6>Ca12>Ca3>CK>Ca24>Ca1水淹(水淹)。显然,Ca6是缓解水分胁迫对亚麻幼苗伤害的最适浓度。此外,本研究还发现,无论是水淹胁迫和干旱胁迫条件下,Ca6和Ca12的影响力与综合评价得分情况是在所有处理组的前2,且两处理组之间的平均得分差值最小,影响力得分在5.79和1.43之间,综合得分在0.02和0.05之间。本研究中的结果仅表明了6和12mmol/L CaCl2的得分和影响力是在所有处理中的前列,在此浓度范围之间可能还有更加适宜的浓度,就目前结果而言,一方面6和12mmol/L CaCl2的综合得分差值不足0.05,6~12mmol/L之间存在比这两个得分更高的可能性较小;另一方面,根据正态分布原理,6和12mmol/L的处理的结果可能是处于正态分布波峰左右两侧处于同一水平位置的结果。具体是否存在某个CaCl2浓度处理的结果处于正态分布波峰顶点的位置,有待于进一步确定,而本研究得到的结果可以应用于亚麻农业生产的理论指导。本研究结果说明,6~12mmol/L CaCl2是缓解水分胁迫对亚麻幼苗伤害的最适浓度,这两个浓度的差异最小得分比价高,进一步说明了缓解两种胁迫最适浓度是同一浓度范围,可以用于亚麻抗旱、抗涝的栽培技术应用和接下来的深入研究。
4 结论
枞形期亚麻根施6~12mmol/L CaCl2是最适缓解水分胁迫的浓度范围,可通过提升CAT活性,降低SOD和POD活性,降低O??含量、H2O2含量和MDA含量,调节PAL活性,缓解水分胁迫对亚麻幼苗的株高、地上部和地下部鲜重、叶片数的抑制,促进亚麻植株形态向正常态生长发育。另外,本研究证明了6mmol/L CaCl2是在水淹和干旱胁迫下对亚麻幼苗缓解效果最好的浓度,进一步证明了缓解两种胁迫最适浓度是同一浓度,为以后的研究提高植物耐旱性、耐水淹性、预防环境胁迫提供了新思路和技术上的支持。
参考文献
[1] Qian Zhao, Shuyao Li, Fu Wang, et al. Transcriptome analysis and physiological response to heat and cold stress in flax (Linum usitatissimum L) at the seedling stage. Environmental and Experimental Botany,2025,229:106076.
[2] 谭金芳. 作物施肥原理与技术. 北京:中国农业大学出版社,2021.
[3] Muhammad S H, Mahantesh M K, Khalil-Ur-Rehman M, et al. Grapevineimmune signaling network in response to drought stress as revealed by transcriptomic analysis. Plant Physiology and Biochemistry,2017,121:187-195.
[4] Mahnaz A, Pooyan M ,Tahereh N, et al. Effect of different primings on germination and growth parameters of rice (Oryza sativa) seeds during drought stress, as compared with silver nanoparticles. Iranian Journal of Seed Research,2018,4(2):121-131.
[5] Zhu Z H, Sami A, Xu Q Q, et al. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE,2021,16(9):e0257236.
[6] 苏芸芸,付菲菲,欧晓彬.外源钙对干旱胁迫下桔梗幼苗生理特性和药用品质的影响.西北植物学报,2024,44(4):551-561.
[7] 郑云海.氯化钙对淹水胁迫下夏玉米生长的调控效应.郑州:河南农业大学,2021.
[8] 李波,刘畅,李红,等.外源氯化钙对‘龙牧807’苜蓿幼苗干旱缓解效应分析.草地学报,2020,28(4):990-997.
[9] 李小玲,华智锐,何娇娇.干旱条件下外源钙对商洛黄芩生理特性的影响.江西农业学报,2019,31(4):31-35.
[10] 刘卫星,张枫叶,张军,等.CaCl2浸种对苗期干旱胁迫下花生生长?生理及产量的影响.山东农业科学,2022,54(3):49-55.
[11] 肖召杰,郑东方,杨秋月,等.不同CaCl2处理对干旱胁迫下花生幼苗生长及生理特性的影响.陕西农业科学,2024,70(8):13-17.
[12] 李盼盼.水淹胁迫下过氧化钙对玉米苗期生理生化特性的影响.泰安:山东农业大学,2022.
[13] 张汉林.外源氯化钙和硝酸钙对渍水胁迫下紫花苜蓿生长和生理特性的影响研究.扬州:扬州大学,2021.
[14] 邓欣,陈信波,龙松华,等.水分胁迫对亚麻苗期生理特性及干物质积累的影响.中国油料作物学报,2015,37(6):846-851.
[15] 蒋虹,张健,王景明,等.《植物生理学实验指导》(第5版)中的几个不足之处.植物生理学报,2020,56(1):11-15.
[16] 施海涛.植物逆境生理学实验指导.北京:科学出版社,2016.
[17] 苏小雨,高桐梅,张鹏钰,等.基于主成分分析及隶属函数法对芝麻苗期耐热性综合评价.作物杂志,2023(4):52-59.
[18] 陈德华.作物栽培学研究实验法.北京:科学出版社,2018.
[19] 何敏敏,王堽,耿贵.钙营养对甜菜幼苗生长和表观光合作用的影响.黑龙江大学自然科学学报,2023,40(3):325-331,379.
[20] 赵海洋,李晓秀,王玉祥,等.氯化钙对干旱胁迫下金莲花幼苗形态特征的影响.农业科技通讯,2018(7):189-193.
[21] 范小玉,陈雷,刘卫星,等.氯化钙浸种对干旱胁迫下花生种子萌发及幼苗生理特性的影响.江苏农业科学,2022,50(8):101-105.
[22] Zhang J, Cheng K, Ma B J, et al. CaCl2 promotes the cross adaptation of Reaumuria trigyna to salt and drought by regulating Na+, ROS accumulation and programmed cell death. Plant Physiology and Biochemistry,2023,195:214-227.
[23] Shah W, Zaman N, Ullah S, et al. Calcium chloride enhances growth and physio-biochemical performance of barley (Hordeum vulgare L.) under drought-induced stress regimes: a future perspective of climate change in the region. Journal of Water and Climate Change,2022,13(9): 3357-3378.
[24] Zhu Z H, Sami A, Xu Q Q, et al. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE,2021,16(9):e0257236.
[25] 樊海潮,顾万荣,杨德光,等.化控剂对东北春玉米茎秆理化特性及抗倒伏的影响.作物学报,2018,44(6):909-919.
[26] 卢昆丽,尹燕枰,王振林,等.施氮期对小麦茎秆木质素合成的影响及其抗倒伏生理机制.作物学报,2014,40(9):1686-1694.
[27] 徐荣琼,张翼飞,杜嘉瑞,等.叶面喷施钙肥对春玉米茎秆抗倒伏特性与产量形成的影响.作物杂志,2024(3):223-230.
[28] Porakala V K, Bineeta M B, Roopa L G, et al. Potential of zinc glycinate and calcium chloride on morphological and yield characters of wheat (Triticum aestivum L). International Journal of Environment and Climate Change,2023,13(10):4031-4037.
[29] 吴儒刚,裴艳婷,张超,等.基于盐胁迫的小麦农艺性状多样性分析及评价.麦类作物学报,2019,39(9):1029-1037.
[30] 赵博慧,张影全,景东林,等.基于多年定点的小麦籽粒质量稳定性研究.中国农业科学,2024,57(9):1833-1844.
[31] 赵康,杨涛,王红刚,等.42个新疆海岛棉品种萌发期耐盐性评价.作物杂志,2022(5):27-33.
[32] 鞠乐,陈培育,牛银亭,等.不同谷子品种(系)萌发期对干旱胁迫的响应及抗旱性评价.江苏农业科学,2023,51(23):40-46.
[33] 苏小雨,高桐梅,张鹏钰,等.基于主成分分析及隶属函数法对芝麻苗期耐热性综合评价.作物杂志,2023(4):52-59.
文章摘自:褚霈宇,韩喜财,王盼,李多,朱浩,金喜军,宋鑫玲,曹洪勋,夏尊民,王晓楠.水分胁迫下CaCl2对亚麻幼苗生长及生理的影响[J/OL].作物杂志.https://link.cnki.net/urlid/11.1808.s.20250523.1007.002。
