摘 要:为了解羽脉山黄麻(Trema levigata)种子的休眠特性,以采自石屏居群(SP)和元谋居群(YM)的种子为材料,探讨不同恒温、变温、赤霉素、光照强度对种子萌发的影响,并对种子的形态、千粒重、含水量进行测量。结果表明,SP和YM的千粒重分别为(1.203±0.005)和(0.945±0.006)g。在恒温条件下,SP仅在30℃下萌发;YM仅在25℃和30℃下萌发;变温对2个居群种子的萌发影响显著且一致,随变温温度增加,发芽率增高,发芽时滞和平均萌发时间减短;赤霉素对2个居群种子的萌发有一定的促进作用,YM尤其显著,说明羽脉山黄麻存在浅型生理休眠;不同光照强度对2个居群种子的萌发影响都不显著,说明羽脉山黄麻属于光中性种子。在30℃/20℃、光照70~140µmol/(m2s)下SP和YM的发芽率分别为86.67%~95.00%和81.67%~95.00%,这是羽脉山黄麻最合适的萌发条件。这为羽脉山黄麻和同属植物造林育苗提供理论依据和技术指导。
关键词:羽脉山黄麻;种子萌发;恒温;变温;赤霉素;光照
山黄麻属(Trema)植物是热带、亚热带地区的先锋树种,在经济、生态和药用方面都有重要的价值[1]–[2]。本属植物韧皮纤维发达,品质优良,可作为造纸、人造棉的原料[3];其植株生长迅速,萌生能力强,根系发达,在荒山荒地,土壤贫瘠地区能迅速生长,是治理水土流失的优良速生树种,对生态平衡的维持具有重要功能[3]–[5];另外,其根和叶具有一定的药用价值,植株内含有多种具有药理活性的化学成分[6]。目前,山黄麻属植物在林业上并未得到推广利用,主要原因是种子在自然状态下萌发困难,有研究表明,西双版纳地区的异色山黄麻(T.orientalis)种子在未经人工处理的情况下萌发率为0,且容易腐烂[7];而印度尼西亚巴厘岛的种群,在正常情况下萌发率只能达到38%,且种子的萌发率与种子质量和直径有关,种子越小,萌发率越低[8]。这些研究综合表明,山黄麻属植物的种子在自然条件下萌发率低且萌发不整齐,在进行人工造林和植被恢复前需要通过一定的处理来促进种子的萌发。为了解决山黄麻属植物种子萌发率低的问题,本研究选择2个居群的羽脉山黄麻(T.levigata)为研究对象,探讨不同恒温、变温、赤霉素、光照强度下种子的萌发特性,以期获得最适合的种子萌发条件,为此属植物的造林育苗提供理论支持和种质资源的保藏提供科学依据。
1 材料和方法
1.1 材料
以羽脉山黄麻(Trema levigata)成熟种子为实验材料,种子来自当年自然散布期采集的新鲜果实,从采集得到的果实中挑选深色成熟果实,选择合适孔径的筛网,轻柔的揉搓,将果肉和种子分离,在流水下将种子淘洗干净并置于吸水纸上晾干,将得到的种子通过种子分离机去除空瘪种子后得到实验种子。2个居群分别位于云南省红河哈尼族彝族自治州石屏县(SP)和云南省楚雄彝族自治州元谋县(YM),生境均为干热河谷。
1.2 方法
种子形态指标测定 随机选取20粒种子,在体视显微镜下观察并记录种子的外部形态,包括形状、颜色、种皮网纹和隆起数量等表面特征;将种子沿中轴线剖开,并进一步将胚完整剥出,在体视镜下拍照,倍数为200,参照Martin[9]的The Comparative Internal Morphology of Seeds对种子形态进行描述;随机选取20粒种子,用电子游标卡尺(精度为0.01mm)测量种子的长、宽和厚度;随机选取100粒种子用电子天平(精度为0.00001g)称重,重复10次,求平均值并换算成千粒重。
萌发实验 种子置于1%琼脂培养基的培养皿中,培养皿加盖置于光照培养箱中进行萌发实验。设置7个恒温温度梯度,分别为5℃、10℃、15℃、20℃、25℃、30℃和35℃,光周期为12h,光照强度为14µmol/(m2s)(冷光源);4个变温温度梯度分别为15℃/5℃、20℃/10℃、25℃/15℃、30℃/20℃,高温12h,有光照,光照强度为14µmol/(m2s)(冷光源),低温12h,黑暗。每个梯度设置3个重复,每个重复20粒种子。每周检查1次种子萌发情况,胚根长出2mm为已萌发,第25周结束实验并统计发芽率。发芽率(%)=(萌发种子数/供试种子数)×100%;发芽时滞(d):从发芽实验开始到第1粒种子开始萌发所需要的天数;平均萌发时间(d)=∑nt/∑n,n为第t天的萌发种子数,t为萌发天数。
赤霉素处理 在5℃、10℃、15℃、20℃、25℃、30℃、35℃的萌发温度基础上,每个梯度设置3个重复,每个重复20粒种子,萌发基质为添加200mg/L赤霉素(GA3)的1%琼脂培养基,光周期为12h,光照强度为14µmol/(m2s)(冷光源)。
光照处理 结合温度梯度实验,选取萌发率最高的温度条件进行光照处理。设置4个光照梯度,分别为70、112、140µmol/(m2s)、黑暗,每个梯度设置3个重复,每个重复20粒种子,萌发基质为1%琼脂培养基,高温12h,有光照,低温12h,黑暗;黑暗条件为24h黑暗无光照。
1.3 数据的统计分析
采用Excel 2013进行数据整理和图表绘制;不同处理进行单因素方差分析,LSD法检验处理间差异性(P<0.05),所有统计分析均在R软件环境中进行。
2 结果和分析
2.1 羽脉山黄麻种子形态特征
SP种子褐色,宽圆形,种子两面凹凸不平,明显隆起2~4条脊,平均每面2.33条;胚周缘型,弯曲、长而大且贴近种皮,占种子3/4左右,胚乳位于种子中央,子叶狭长。YM种子褐色,近圆形或宽圆形,种子两面凹凸不平,明显隆起1~3条脊,平均每面1.85条;胚形态与SP一致。YM的千粒重、种子的长、宽、厚均大于SP,2份种子的长短径比接近,2个居群的种子形态差异不显著(表1,图1)。
表1 2个居群羽脉山黄麻种子形态特征

SP:石屏居群;YM:元谋居群;同列数据后不同字母表示显著差异(P<0.05)。下同
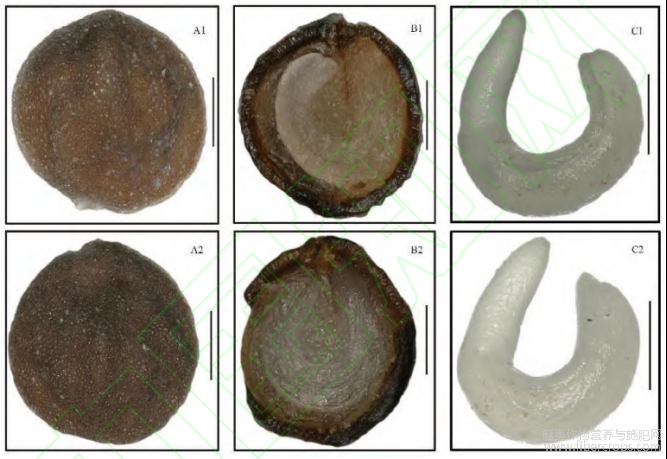
图1 羽脉山黄麻SP居群(A1~C1)和YM居群(A2~C2)的种子形态
A:形态;B:横切面;C:胚。标尺=500μm。
2.2 恒温对种子萌发的影响
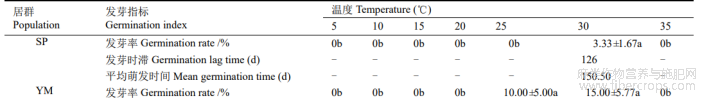
在恒温条件下,SP种子在5℃~25℃、35℃下都不萌发,仅在30℃下萌发,发芽率为3.33%,发芽时滞为126d,平均萌发时间为150.50d,与其他条件差异显著。YM种子5℃~20℃和35℃下都不萌发,仅在25℃和30℃下萌发,30℃下的发芽率高于25℃,但无显著差异;30℃下的发芽时滞、平均萌发时间比25℃下短(表2)。
表2 恒温对羽脉山黄麻种子萌发的影响
![]()
-: 种子未萌发,无数据。下同
2.3 变温对种子萌发的影响
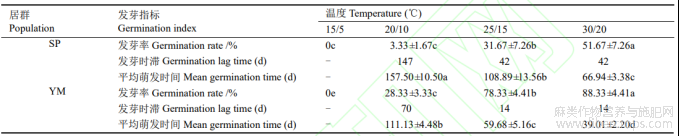
由表3可见,SP和YM种子在15℃/5℃下不萌发,在20℃/10℃、25℃/15℃、30℃/20℃下萌发,且各萌发条件间差异显著,随变温温度增加,发芽率增高,平均萌发时间减短。SP在30℃/20℃下发芽率最高,平均萌发时间最短;20℃/10℃下发芽率最低,平均萌发时间最长;25℃/15℃和30℃/20℃下发芽时滞最短,较20℃/10℃下缩短105d。YM种子在30℃/20℃下发芽率最高,平均萌发时间最短;20℃/10℃下发芽率最低,平均萌发时间最长;25℃/15℃和30℃/20℃下发芽时滞最短,较20℃/10℃下缩短56d。
表3 变温对羽脉山黄麻种子萌发的影响
2.4 赤霉素对种子萌发的影响
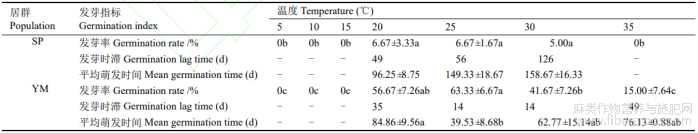
在高于20℃(含)的恒温条件下,赤霉素对羽脉山黄麻种子萌发有一定促进作用(表4)。SP种子在5℃、10℃、15℃和35℃下不萌发,仅在20℃、25℃和30℃下萌发,发芽率为5.00%~6.67%,但差异均不显著;随温度升高,发芽时滞和平均萌发时间变长,20℃下发芽时滞为49d,较30℃缩短77d,平均萌发时间最短。YM种子在5℃、10℃和15℃下不萌发,20℃~35℃下萌发,其中25℃发芽率最高,平均萌发时间最短;35℃发芽率最低,发芽时滞最长;25℃、30℃和35℃间的发芽率差异显著。
表4 赤霉素对羽脉山黄麻种子萌发的影响
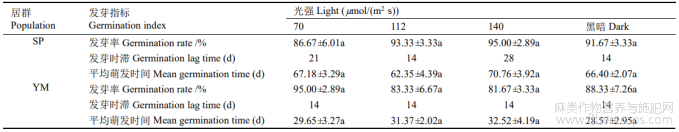
2.5 光照对种子萌发的影响
前期实验表明,30℃/20℃为羽脉山黄麻种子萌发的最佳条件。在该温度下进行光照处理,SP和YM种子在光照和黑暗条件下均能萌发,且光照和黑暗间的发芽率和平均萌发时间无显著差异。SP发芽率为86.67%~95.00%,在140µmol/(m2s)的光照强度下的发芽率最高;70µmol/(m2s)下最低;112µmol/(m2s)的平均萌发时间最短。YM发芽率为81.67%~95.00%,平均萌发时间为28.57~32.52d,以70µmol/(m2s)的光照强度下的发芽率最高,140µmol/(m2s)下最低;黑暗下平均萌发时间最短;光照和黑暗的发芽时滞相同(表5)。
表5 光照强度对羽脉山黄麻种子萌发的影响
3 讨论和结论
具有生命力的种子在适宜的萌发条件下不能萌发的现象称为种子休眠。种子休眠是植物在长期系统发育过程中对不良环境条件形成的适应性,是调节种子萌发最佳时间和空间分布的有效方法,对植物本身的繁衍具有重要意义[10]。
引起种子休眠的原因有很多种,目前,在研究领域较为广泛接受和应用的种子休眠分类方法为Baskin等[11]提出的种子休眠的分类方法,即分为生理休眠、形态休眠、形态生理休眠、物理休眠和综合休眠5种类型。种子休眠的破除包括物理方法(机械处理、温度处理、射线、超声波、电场和磁场处理、干藏、干湿处理)、化学方法(激素处理、无机化学试剂处理、有机化学药剂处理)、生物处理、综合处理等。造成植物种子休眠的原因往往是由多种因素引起的,综合采取多种措施,可在一定程度上打破休眠,促进种子萌发[12]。
3.1 温度对种子萌发的影响
温度是影响种子萌发的重要因素之一。对温度敏感的种子,适宜的温度可以促进种子的萌发,而过高或过低的温度则会抑制种子的萌发[13]。对于某些物种,变温条件是打破种子休眠,促进种子萌发的有效手段[16]–[17]。变化的温度可以使种皮胀缩,从而有利于水分和气体进入种子内部,促进种子内外温差的形成,增加气体交换,有利于氧气的供应,促进呼吸作用,增强与发芽有关的酶活性;在变温条件下,高温时生化过程和呼吸都较为旺盛,有利于贮藏物质转化为可溶性物质,低温时呼吸减弱,物质消耗减少,有利于胚的生长[18]–[21]。通过变温处理,可以模拟自然环境中的温度变化,为种子提供更接近自然的生长条件,从而促进种子的萌发。本研究表明SP在恒温条件下只在30℃下萌发,YM只在25℃、30℃下萌发,说明羽脉山黄麻对温度敏感,但在该条件下,发芽率低且发芽时滞和平均萌发时间长。在变温条件下,SP和YM表现出一致性,在20℃/10℃、25℃/15℃和30℃/20℃下均能萌发,随变温温度升高发芽率增高,发芽时滞和平均萌发时间变短。实验结果表明变温条件优于恒温,且30℃/20℃为最优萌发温度。
3.2 赤霉素对种子萌发的影响
赤霉素是一种具有多种生理功能的植物生长激素,不仅促进植物生长发育和对各种胁迫的应答,还能打破一些植物的种子休眠,促进细胞伸长和分裂以及调节种子的生理状态,对种子萌发产生促进作用[22]–[23]。本研究表明在赤霉素处理的条件下,SP和YM的发芽率高于同温度下无赤霉素的处理,发芽时滞和平均萌发时间显著变短,YM尤其明显,因此通过使用赤霉素能够解除羽脉山黄麻种子的休眠,确定其休眠类型为生理休眠[11]。进一步的分析表明未添加赤霉素处理下SP仅在30℃下萌发,赤霉素处理后能在20℃、25℃和30℃萌发;未添加赤霉素处理下YM仅在25℃和30℃下萌发,赤霉素处理后能在20℃、25℃、30℃和35℃下萌发。赤霉素的处理不仅在一定程度下解除了种子的休眠,提高了发芽率,也使种子能在更低的温度下萌发,拓宽了种子的萌发温度范围,因此羽脉山黄麻种子的休眠类型进一步细分为浅型生理性休眠中的第二种类型(type 2 non-deep PD)[11]。综合研究表明赤霉素处理可以在一定恒温下有效打破羽脉山黄麻种子的休眠并缩短种子萌发周期。
研究进一步表明非休眠的种子具有明显的萌发温度偏好,SP和YM经过赤霉素处理后萌发温度高于20℃,这与生境的气候密切相关。种子解除休眠后,在适宜的环境中萌发能确保幼苗免受高温或低温等逆境伤害,且萌发温度的选择常与降雨发生的时间密切相关[11]。2个种子采集地均为干热河谷,生境相似。石屏县年均温度约为18℃,年均降水量为775~1148mm,主要集中在5月—7月[24]。元谋县年均气温为21.9℃,年均降水量仅有616mm,雨热同期,全年90%的降水集中在6月—10月(雨季),期间月均温高于20℃[25]。羽脉山黄麻种子的萌发温度与雨季温度一致,避免种子在其他时间萌发后,幼苗受到干旱逆境的伤害。从生态学的观点看,萌发行为的温度偏好性可能是植物长期适应其生境而进化的结果。温度不仅能调控种子休眠状态的变化,也能直接影响种子的萌发行为[11]。如经冷层积(4℃)或GA3溶液处理后,落地梅(Lysimachia paridiformis)种子能在20℃/10℃下顺利萌发,但不能在10℃和20℃等温度下顺利萌发[26]。
3.3 光照对种子萌发的影响
光照是影响植物种子萌发过程中重要的生态环境因子之一[27]。不同植物的种子在萌发过程对光的需求有所不同,光照对需光性种子的萌发有促进作用,对忌光性种子的萌发有抑制作用,而对光中性种子无影响[30]。本研究表明在不同光照强度和黑暗条件下羽脉山黄麻种子的发芽率、平均萌发时间之间差异不显著,说明光照不是羽脉山黄麻萌发的必要因素,羽脉山黄麻种子为光中性种子,与王直军等[7]的研究结果不同。另外,有研究表明当种子重量超过0.1mg时,种子萌发对光照的需求性就不明显[33],本实验中SP千粒重为1.203g,YM为0.945g,与其相似。
综上,从恒温、变温、赤霉素、光照等条件探索羽脉山黄麻种子的萌发特性,结果表明,2个居群种子的萌发特性较为一致,都存在浅型生理休眠,赤霉素能够打破休眠,同时变温条件下的发芽率显著高于恒温和赤霉素处理;变温30℃/20℃下,光照70~140µmol/(m2s)是羽脉山黄麻最适合的萌发条件,这可作为羽脉山黄麻和同属山黄麻植物造林育苗的参考和依据。
参考文献
[1]LIU G, YAO C H, CHEN H Y. Study on biology characteristic and exploitation potential of Trema levigata Hand. [J]. Hubei For Sci Technol, 2006, 35(4): 29–34.[刘刚,姚崇怀,陈红艺.羽脉山黄麻的生物学特性及其开发潜力的研究[J].湖北林业科技,2006,35(4):29–34.doi:10.3969/j.issn.1004-3020.2006.04.009.]
[2] YANG C B. Drought-tolerant “pioneer tree species”: Trema orientalis [J]. Yunnan For, 2013, 34(2): 64.[杨超本.耐旱的“先锋树种”——山黄麻[J].云南林业,2013,34(2):64.]
[3] HAN S Z, CHEN K L, YIN Y D, et al. Evaluation of Trema Orientalis as a papermaking raw material [J]. China Pulp Paper, 2019, 38(11): 32–36.[韩绍中,陈克利,尹玉冬,等.山黄麻作为造纸原料的基础评价[J].中国造纸,2019,38(11):32–36.doi:10.11980/j.issn.0254-508X.2019.11.005.]
[4] YANG C B, DENG G H, YANG C J, et al. Study on afforestation technology of Trema Orientalis in dry-hot valley of South China [J]. Pract For Technol, 2014(4): 12–14.[杨超本,邓光华,杨祟健,等.南方干热河谷地区山黄麻造林技术研究[J].林业实用技术,2014(4):12–14.doi:10.13456/j.cnki.lykt.2014.04.004.]
[5] HE M S, ZHANG E X. Trema tomentosa seedling and afforestation technology [J]. For Sci Technol, 2022(6): 98–99.[何明阐,张恩向.山黄麻育苗造林技术[J].林业科技通讯,2022(6):98–99.doi:10.13456/j.cnki.lykt.2021.10.13.0002.]
[6] LI Y, ZHANG D M, YU S S. Studies on chemical constituents and pharmacological activities of Trema [J]. J Chin Med Mat, 2003, 26(11): 833–838.[李媛,张东明,庾石山.山黄麻属植物的化学成分和药理活性研究[J].中药材,2003,26(11):833–838.doi:10.3321/j.issn:1001-4454.2003.11.030.]
[7] WANG Z J, CAO M, LI G F, et al. Trema orientalis seeds dispersed by birds and its ecological role [J]. Zool Res, 2002, 23(3): 214–219.[王直军,曹敏,李国锋,等.鸟类对山黄麻种子的传播及其生态作用[J].动物学研究,2002,23(3):214–219.doi:10.3321/j.issn:0254-5853.2002.03.006.]
[8] NUGRAHENI Y M M A, YUNIARTI N. The impact of drying time and storage room on the viability of trema (Trema orientalis Linn. Blume) seed [J]. IOP Conf Ser Earth Environ Sci, 2022, 1109(1): 012097. doi: 10.1088/1755-1315/1109/1/012097.
[9] MARTIN A C. The comparative internal morphology of seeds [J]. Am Midl Nat, 1946, 36(3): 513–660. doi: 10.2307/2421457.
[10] ZHANG H S, HU J. Seed Science [M]. 2nd ed. Beijing: Science Press, 2015: 106–107.[张红生,胡晋.种子学[M].第2版.北京:科学出版社,2015:106–107.]
[11] BASKIN C C, BASKIN J M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination [M]. 2nd ed. San Diego: Academic Press, 2014.
[12] FU N, SONG H, WANG S J, et al. Research progress on seed dormancy and breaking methods [J]. J Anhui Agric Sci, 2018, 46(24): 10–12, 15.[付楠,宋慧,王淑君,等.种子的休眠与破除研究进展[J].安徽农业科学,2018,46(24):10–12,15.doi:10.13989/j.cnki.0517-6611.2018.24.003.]
[13] SHI J Y, SUN S J, MA W, et al. Study on germination characteristics and antioxidant physiology of Siberian wildrye seeds under different temperatureconditions [J]. Acta Agrest Sin, 2024, 32(10): 1–12. [石金玉,孙守江,马馼,等.不同温度条件下老芒麦种子萌发特性和抗氧化生理研究[J].草地学报,2024,32(10):1–12.doi:10.11733/j.issn.1007-0435.2024.10.015.
[14] GENG B Y, QIN Y B, YANG L E, et al. Response of seed germination of the alpine endangered species Rhodiola crenulata to temperature in different years [J]. Pratacult Sci, 2024, 41(8): 1924–1933.[耿博阳,覃滢卜,杨丽娥,等.不同采集年份大花红景天种子萌发对温度的响应[J].草业科学,2024,41(8):1924–1933.doi:10.11829/j.issn.1001-0629.2023-0250.]
[15] WANG Y X, ZHANG X, WANG L, et al. Effects of seed size and temperature changes on seed germination characteristics of wild medicinal plants in desert region [J]. Seed Sci Technol, 2024, 42(11): 46–50.[王奕璇,张雪,王磊,等.种子大小和温度变化对荒漠区野生药用植物种子萌发特性的影响[J].种子科技,2024,42(11):46–50.doi:10.19904/j.cnki.cn14-1160/s.2024.11.014.]
[16] BASKIN J M, BASKIN C C. A classification system for seed dormancy [J]. Seed Sci Res, 2004, 14(1): 1–16. doi: 10.1079/SSR2003150.
[17] KUMAR B, GUPTA E, MALI H, et al. Constant and alternating temperature effects on seed germination potential in Artemisia annua L. [J]. J Crop Improv, 2013, 27(6): 636–642. doi: 10.1080/15427528.2013.832458.
[18] HU X W, HUANG X H, WANG Y R. Hormonal and temperature regulation of seed dormancy and germination in Leymus chinensis [J]. Plant Growth Regul, 2012, 67(2): 199–207. doi: 10.1007/s10725-012-9677-3.
[19] OZDEN E, LIGHT M E, DEMIR I. Alternating temperatures increase germination and emergence in relation to endogenous hormones and enzyme activities in aubergine seeds [J]. S Afr J Bot, 2021, 139: 130–139. doi: 10.1016/j.sajb.2021.02.015.
[20] WANG J T, YANG J, WANG X T. The effect of the changing temperature treatment on the germination of three Gramineae grass seeds in the northern Xizang Plateau [J]. J Plateau Agric, 2019, 3(6): 670–674.[王江涛,杨军,王向涛.变温处理对藏北高原3种禾本科牧草种子萌发的影响[J].高原农业,2019,3(6):670–674.doi:10.19707/j.cnki.jpa.2019.06.011.]
[21] WANG X, XI Z, WU F, et al. Study on seed germination characteristics of Ensete glaucum [J]. Seed, 2023, 42(2): 128–132.[王熙,西战,吴菲,等.象腿蕉种子萌发特性研究[J].种子,2023,42(2):128–132.doi:10.16590/j.cnki.1001-4705.2023.02.128.]
[22] YANG Y F, LUO L K, DING C, et al. Effects of gibberellin on seed germination and seedling growth of Parrotia subaequalis, a rare and endangered plant species [J]. Chin J Ecol, 2024, 43(7): 2026–2032. [阳艳芳,罗来开,丁聪,等.赤霉素对珍稀濒危植物银缕梅种子萌发及幼苗生长的影响[J].生态学杂志,2024,43(7):2026–2032.doi:10.13292/j.1000-4890.202407.002.]
[23] SONG Z F, ZHANG Z M, LI M J, et al. Effects of abscisic acid and gibberellin on growth and post-ripening process of peanut [J]. J Peanut Sci, 2024, 53(2): 10–19. [宋兆锋,张志民,李美君,等.赤霉素和脱落酸对花生生长与后熟进程的影响[J].花生学报,2024,53(2):10–19.doi:10.14001/j.issn.1002-4093.2024.02.002.]
[24] LI Y, ZHANG C F, ZHANG L X, et al. Investigation and diversity analysis of medicinal plant resources in Shiping County, Yunnan Province [J]. J Chin Med Mater, 2024, 47(1): 39–44. [李瑶,张翠峰,张丽霞,等.云南省石屏县药用植物资源调查与多样性分析[J].中药材,2024,47(1):39–44.doi:10.13863/j.issn1001-4454.2024.01.007.]
[25] HE C. Study on the relationship between vegetation and roil physical and chemical properties of typical eroded gully in Yuanmou Dry-hot Valley region [D]. Nanchong: China West Normal University, 2021. [何聪.元谋干热河谷典型侵蚀沟植被与土壤理化性质关系研究[D].南充:西华师范大学,2021.doi:10.27859/d.cnki.gxhsf.2021.000039.]
[26] TIAN M H, ZHANG Y Y, YANG Y Q, et al. Dormant characteristics and germination behavior in ornamental Lysimachia paridiformis seeds [J]. Seed, 2016, 35(9): 32–36.[田美华,张有义,杨永清,等.非深度生理休眠的落地梅种子的萌发行为[J].种子,2016,35(9):32–36.doi:10.16590/j.cnki.1001-4705.2016.09.032.]
[27] LI C, LU T, LIU Y, et al. Effects of different treatments on seed germination of Enkianthus chinensis Franch. [J]. N Hort, 2020(22): 72–78. [李超,卢婷,刘阳,等.不同处理对灯笼树种子萌发的影响[J].北方园艺,2020(22):72–78.doi:10.11937/bfyy.20194815.]
[28] JIANG L, PENG M Z, TIAN C Y. Effects of temperature, salinity and light on seed germination characteristics of desert plant Kalidium foliatum [J]. Seed, 2022, 41(12): 14–18.[姜黎,彭钼植,田长彦.温度、盐分和光照对荒漠植物盐爪爪种子萌发特征的影响[J].种子,2022,41(12):14–18.doi:10.16590/j.cnki.1001-4705.2022.12.014.]
[29] LIANG R F, WU Z N, LI Z Y, et al. Effects of low-temperature and light on seed germination of Leymus chinensis [J]. Chin J Grassland, 2021, 43(12): 33–39. [梁润芳,武自念,李志勇,等.低温和光照对羊草种子萌发的影响[J].中国草地学报,2021,43(12):33–39.doi:10.16742/j.zgcdxb.20200202.]
[30] DONG L, XU A K, WANG Y Z, et al. Effects of different treatments on seed germination of wild golden chrysanthemum [J]. Mol Plant Breed, 2019, 17(23): 7879–7885. [董兰,徐安凯,王英哲,等.不同处理对长白山野生黄金菊种子萌发的影响[J].分子植物育种,2019,17(23):7879–7885.doi:10.13271/j.mpb.017.007879.]
[31] JIAO D Y, TAN Y H, TANG S X, et al. Ecological characteristics of germination of Plukenetia volubilis seeds [J]. J Trop Subtrop Bot, 2011, 19(6): 529–535.[焦冬英,谭运红,唐寿贤,等.星油藤种子萌发的生态学特性研究[J].热带亚热带植物学报, 2011, 19(6): 529–535. doi: 10.3969/j.issn.1005-3395.2011.06.007.]
[32] ZOU W, CHEN Z, JIANG Y Q, et al. Responses of Pinus yunnanensis seed germination to temperature, light and GA3 [J]. J Sichuan For Sci Technol, 2023, 44(1): 84−90. [邹雯,陈卓,蒋云清,等.云南松种子萌发对温度、光照和GA3的响应[J].四川林业科技,2023,44(1):84–90.doi:10.12172/202203180001.]
[33] GRIME J P, MASON G, CURTIS A V, et al. A comparative study of germination characteristics in a local flora [J]. J Ecol, 1981, 69(3): 1017–1059. doi: 10.2307/2259651.
文章摘自:丁艳蓉,王怡然,郑艳玲,等.两个居群羽脉山黄麻种子萌发特性研究[J/OL].热带亚热带植物学报,1-8[2025-06-04].http://kns.cnki.net/kcms/detail/44.1374.Q.20250519.1113.004.html.
